Konjugacijski prenos plazmidov pri bakteriji Bacillus subtilis
Uvod
Konjugacija je najbolj pogost mehanizem za horizontalni prenos genov in kot taka tudi najbolj zaslužna za širjenje genov za virulenco, toksine in odpornost proti antibiotikom. Posledično je eden izmed ciljev sodobnih raziskav na področju molekularne biologije razumevanje mehanizmov, po katerih konjugacija poteka. Medtem ko so procesi njene regulacije pri Gramnegativnih bakterijah, katerih predstavnica je E. Coli in njen plazmid IncF, že dokaj dobro raziskani, pri Grampozitivnih bakterijah ni tako. Nova odkritja na področju konjugacijskega prenosa plazmidov pri G+ bakteriji Bacillus subtilis bi lahko pripomogla k zmanjšanju te vrzeli.
Konjugacija pri Bacillus subtilis
Pri procesu konjugacije sodeluje ogromno število različnih proteinov, nastanek katerih mora celica skrbno regulirati. Genski zapis za te konjugacijske proteine je lahko bodisi integriran v bakterijski kromosom ali pa je ohranjen na plazmidu, kakršen je tudi plazmid pLS20, najden v bakteriji Bacillus subtilis. Začetne raziskave pri tem bacilu so pokazale, da mora biti potek konjugacije pri njem strogo reguliran, saj je bila konjugacija ves čas blokirana razen v ozkem časovnem intervalu na koncu eksponentne rasti celic.
RcopLS20 deluje kot represor konjugacijskega operona
V plazmidu pLS20 so nekateri geni izkazovali podobnost z že znanimi konjugacijskimi geni. Izkazalo se je, da ti geni spadajo pod isti operon, ki zajema gene od 28 do 74. Sekvenciranje ter in silico analiza plazmida sta nakazali, da bi transkripcijo na tem operonu lahko reguliral protein RcopLS20, ki ga zapisuje gen 27c, saj se gen nahaja pred operonom in mu je nasprotno usmerjen, njegovo zaporedje pa je podobno zaporedju za transkripcijske regulatorje Xre, ki imajo heliks-zavoj-heliks domeno. Z eksperimenti, kjer so opazovali izražanje RNA konjugacijskih genov pri različnih koncentracijah domnevnega regulatorja, so potrdili represorsko funkcijo RcopLS20.
Sestava operatorske regije konjugacijskega operona
V regiji med genoma 27c (represor RcopLS20) in 28 (konjugacijski operon) se nahajata dva promotorja: močan promotor Pc za prepisovanje konjugacijskega operona in šibek promotor Pr za prepisovanje RcopLS20. Promotorja sta nasprotno usmerjena in se prekrivata, kar pomeni, da bi se brez drugih dejavnikov (beri: regulatorjev) RNA polimeraza vezala samo na močnejši Pc. V okolici promotorjev so našli 10 polj z 8 bp dolgim zaporedjem CAGTGAAA, na katera se lahko veže RcopLS20. Polja se nahajajo v dveh regijah, ki nosita funkcijo operatorjev. Prvi operator (O1), ki vsebuje 4 Rco vezavna mesta, se nahaja na območju med genom 28 in promotorjema. Drugi operator (O2) s 6 RcopLS20 vezavnimi mesti pa se nahaja 75 bp stran od O1 in se prekriva s promotorjema. Operatorja sta med sabo nasprotno usmerjena, medtem ko so Rco vezavna mesta v posameznem operatorju enako orientirana.
Mehanizem RcopLS20 represije vključuje nastanek DNA zanke
Položaj in usmerjenost operatorjev sta sprožila sum, da RcopLS20 z vezavo na operator povzroči nastanek DNA zanke in s tem inhibira promotor Pc. Čeprav je za nastanek zanke potrebnih najmanj 90 bp DNA, razdalja med operatorjema pa je 75 bp, bi bila zanka v tem primeru vseeno možna, saj je vijačnica med operatorjema predhodno upognjena. Raziskave so pokazale tudi, da protein RcopLS20 v raztopini tvori tetramere. To dejstvo je skupaj z rezultati testa zamika elektroforezne mobilnosti za regulatorno regijo z vezanim RcopLS20 omogočilo določitev modela vezave RcopLS20. Tetramer RcopLS20 se veže na posamezni operator. Dva tetramera se nato povežeta (tvorita oktamer) in tako ustvarita DNA zanko. V tej konformaciji regulatorne regije je promotor Pc blokiran, kar omogoča vezavo RNA polimeraze na promotor Pr.
Prekinitev represije povzroči RappLS20
Kaseta rap-phr
Da konjugacija lahko poteče, mora priti do prekinitve represije, to pa se v tem primeru zgodi preko proteina RappLS20. Ta je del družine proteinov RRNPP, poimenovano po značilnih predstavnikih Rap, Rgg, NprR, PlcR in PrgX. Geni za proteine RRNPP, rap, se običajno prepisujejo istočasno kot geni phr, ki zapisujejo ustrezen signalen peptid Phr*pLS20, o katerem bomo izvedeli več kasneje. Kromosom B. subtilis vsebuje več rap-phr kaset, ki služijo zaustavljanju določenih razvojnih procesov kot je sporulacija, kompetenca in proizvodnja razgradnih encimov. Umetno povečano izražanje teh kaset na plazmidu pLS20 v B. subtilis ni vplivalo na razvojne procese, je pa močno vplivalo na konjugacijski proces, povečala se je največja raven konjugacije, povečalo se je izražanje konjugaciskih genov in razširilo se je časovno obdobje, v katerem je lahko uspešno potekala konjugacija. Pri odsotnosti kaset rap-phr pa je raven konjugacij močno upadla. Ti rezultati nakazujejo, da so RcopLS20, RcopLS20 in Phr*pLS20 edini proteini zapisani napLS20, ki so vpleteni v regulacijo konjugacijskih genov.
RRNPP proteini
RRNPP proteini so sestavljeni večinoma iz alfa heliksov in vsebujejo veliko C-terminalno domeno, ki interagira z ustreznim signalnim peptidom ter povzroči konformacijske spremembe in N-terminalno domeno, ki vsebuje efektorsko domeno sestavljeno iz treh alfa vijačnic, ki interagira s tarčno molekulo, svojo regulatorno vlogo pa lahko opravlja na različne načine. Prvi način je že omenjeni motiv heliks-zavoj-heliks, drugi je fosfatazna aktivnost, po kateri se imenujejo proteini Rap (Response regulator Aspartate Phosphatase) in tretji način, vezava efektorske domene na ustrezajoči efektorski protein, s čimer posredno ali neposredno zavre aktivnost efektorskega proteina.
Delovanje RappLS20
Efektorska domena RappLS20 deluje na tretji način, veže se neposredno na tetramer RcopLS20, kar povzroči, da se le ta odcepi od operatorjev. To je bilo ugotovljeno z analizo SAXS in velikostno izključitvene kromatografijo, poskusi sedimentacijske hitrosti pa so pokazali na tvorbo strukture, ki je ustrezala molekularni teži kompleksa RappLS20 dimera in RappLS20 tetramera. Z EMSA analizo je bilo ugotovljeno tudi, da se dimer RappLS20 preferenčno veže na tetramere že vezane v DNA zanke. DNA-zanke se razdrejo in se prekine represija.
Vpliv peptida Phr*pLS20
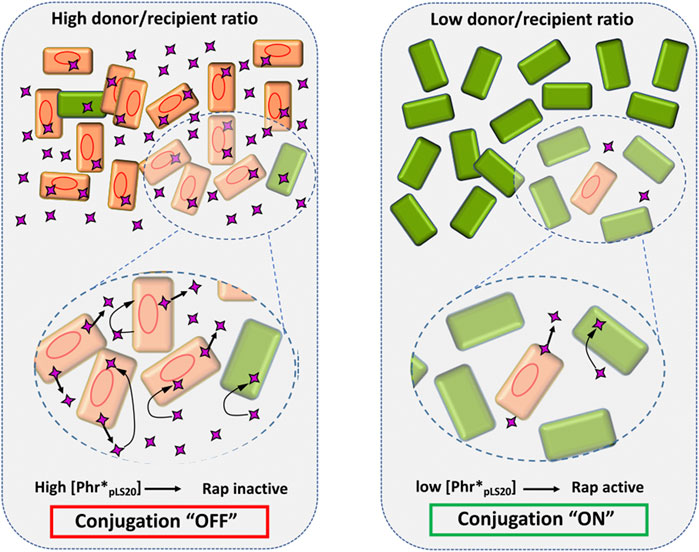
Bakterije uporabljajo kemične signale, imenovane feromoni, za usklajevanje izražanja genov in vedenja celic v skupnosti. Feromoni nekaterih grampozitivnih bakterij, kot je Bacillus subtilis, so kratki linearni peptidi, ki jih celica izloča, nato pa jih zaznajo citoplazemski feromon receptorji iz družine RRNPP. Peptid Phr*pLS20 kodira majhen odprt bralni okvir (44 aminokislin), po sekreciji pa ga cepi serinska proteaza, da nastane zrela aktivna oblika peptida. Tako kot pri drugih RRNPP proteinih in signalnih peptidih, tudi Phr*pLS20 interagira direktno s C-končno domeno RappLS20. Analize kažejo, da RappLS20 v raztopini večinoma tvori dimere, pri višjih koncentracijah pa ima tudi sposobnost tvoriti tetramere. Medtem pa ob prisotnosti Phr*pLS20 RappLS20 skoraj izključno tvori tetramere. RappLS20 je sestavljen iz velike C-končne domene iz 14 α vijačnic ter manjše N-končne domene iz 3 α vijačnic. Struktura z vezanim peptidom je pokazala, da se Phr*pLS20 veže v votlino C-končne domene, kar povzroči, da se N-končna domena premakne navzven. Ta položaj bolj ustreza tvorbi tetramer, pri čemer se dva dimera medsebojno povežeta prek N-končne domene, kar vodi do nastanka tako imenovane stopalo-stopalo konfiguracije. Ker so N-končne domene RappLS20 zasedene, ta ne more interagirati z RcopLS20, torej Phr*pLS20 inhibira antirepresorsko aktivnost RappLS20 in s tem zavira izražanje konjugacijskih genov. Skupno gledano aktivacijsko stanje pLS20 konjugacijskih genov regulira koncentracija peptida Phr*pLS20. Ko večina celic populacije vsebuje plazmid, bo koncentracija peptida visoka in konjugacija inhibirana. Konjugacijski geni bodo aktivni, ko bo koncentracija Phr*pLS20 nizka, kar se zgodi, če so donorske celice v manjšini in potencialne prejemne celice v večini. Veliko število celic, ki lahko sprejmejo peptid, zniža koncentracijo Phr*pLS20, saj se relativno število donorskih celic zmanjša, poleg tega pa prejemne celice aktivno adsorbirajo signalni peptid. Slika1. Kaj se zgodi s kompleksom Rap-Phr ni znano, najbrž so kompleksi hitro razgrajeni ali pa obstaja mehanizem za recikliranje, znano pa je, da je promotor Prap konstituiven, torej se Rap in Phr stalno sintetizirata.
{kind=link}
Konjugacija in kompetenca
Kompetenca je začasno stanje celic, v katerem so celice sposobne sprejeti DNA iz okolja in jo integrirati v svoj genom s homologno rekombinacijo. Konjugacija in kompetenca sta si podobna v tem, da gre pri obeh procesih za prenos enoverižne DNA(ssDNA), le da v nasprotnih smereh. Sočasen vnos in iznos regij ssDNA je za celico lahko nekompatibilno, saj se lahko ta transportna mehanizma medsebojno tekmujeta in se motita. Prav tako lahko rekombinantni encimi sintetizirani med fazo kompetence delujejo na ssDNA konjugativnega elementa. Dokazano je bilo, da sta konjugacija in kompetenca nekompatibilna procesa in obstajata vsaj 2 mehanizma, ki z recipročno inhibicijo skrbita, da se poti konjugacije in kompetence ne aktivirata sočasno.
Zaključek
V primerjavi z G- bakterijami sta konjugacija in regulacija konjugacijskih genov pri G+ bakterijah slabše poznana. Pri Bacillus subtilis je razen regulacije plazmida pLS20 znan tudi mehanizem ICEBs1 (integrativni konjugativni elementi), imenovan tudi konjugativni transpozoni. Nahajajo se na kromosomih in se lahko izrežejo ter zvijejo, nato pa prenesejo v druge celice. Gene ICEBs1 inhibira ImmR , ki je analog RcopLS20, antirepresor pa je v tem primeru RRNPP protein RapI. Celoten mehanizem delovanja RapI znan, je pa za njegovo delovanje potrebna še proteaza, ki razgradi ImmR. Ker je ImmR razgrajen, je konjugacija ICEBs1 ireverzibilen proces, medtem ko pri pLS20 peptid Phr nudi fleksibilnost in reverzibilnost, saj se RcopLS20 po tem ko Phr*pLS20 inaktivira RappLS20 sprosti in spet opravlja vlogo represorja Proces konjugacije je energijsko zahteven proces in ima velik vpliv na celice, zato mora biti proces pod strogo kontrolo, ki dovoli aktivacijo le pri optimalnih pogojih. V praksi je konjugacija plazmida pLS20 iz Bacillus subtilis omejena na čas med zgodnjo in pozno eksponentno rastjo. Eksperimenti izvedeni v preglednem članku so bili izvedeni na tekočih kulturah, ki so bile stresane, kar zagotavlja homogenizacijo signalnih peptidov. Ti pogoji so drugačni od naravnih, kjer se v različnih delih kulture ustvarijo koncentracijski gradienti, zato so potrebne še nadaljnje študije mehanizmov konjugacije.
Viri
Meijer, Wilfried J. J., idr. „Multiple Layered Control of the Conjugation Process of the Bacillus subtilis Plasmid pLS20“. Frontiers in Molecular Biosciences, let. 8, marec 2021, str. 648468. DOI.org (Crossref), https://doi.org/10.3389/fmolb.2021.648468.
Ramachandran, Gayetri, idr. „A Complex Genetic Switch Involving Overlapping Divergent Promoters and DNA Looping Regulates Expression of Conjugation Genes of a Gram-Positive Plasmid“. PLoS Genetics, uredil Danielle A. Garsin, let. 10, št. 10, oktober 2014, str. e1004733. DOI.org (Crossref), https://doi.org/10.1371/journal.pgen.1004733.
Singh, Praveen K., idr. „Mobility of the Native Bacillus Subtilis Conjugative Plasmid pLS20 Is Regulated by Intercellular Signaling“. PLoS Genetics, uredil Josep Casadesús, let. 9, št. 10, oktober 2013, str. e1003892. DOI.org (Crossref), https://doi.org/10.1371/journal.pgen.1003892.