Rastlinski virusi: Difference between revisions
No edit summary |
No edit summary |
||
| (3 intermediate revisions by the same user not shown) | |||
| Line 10: | Line 10: | ||
Tobamovirusi so edini rastlinski virusi paličaste oblike z enodelnim genomom. Virus se prenaša prek rastlinskih sokov in včasih tudi prek okuženih semen. | Tobamovirusi so edini rastlinski virusi paličaste oblike z enodelnim genomom. Virus se prenaša prek rastlinskih sokov in včasih tudi prek okuženih semen. | ||
Genom tega virusa obsega 4 odprte bralne okvirje in je dolg približno 6.0 do 6.5 kb v obliki linearne ssRNA. Na 5' koncu ima metilirano kapo (m7G5’pppG), 3' konec pa ima strukturo podobno tRNA. Zanimivo je to, da 2 od 4 odprtih bralnih okvirjev kodirata za en protein zaradi puščajočega UAG stop kodona, njun produkt pa je replikaza z metilacijsko in helikazno podenoto, hkrati pa mala enota deluje kot zaviralec utišanja RNA, ki je obrambni mehanizem gostitelja. Movement protein (MP) pomaga pri sestavljanju novega viriona, v poznejših fazah replikacije, zapis zanj pa najdemo v subgenomski RNA. Prav tako za CP, kapsidni protein. | Genom tega virusa obsega 4 odprte bralne okvirje in je dolg približno 6.0 do 6.5 kb v obliki linearne ssRNA. Na 5' koncu ima metilirano kapo (m7G5’pppG), 3' konec pa ima strukturo podobno tRNA. Zanimivo je to, da 2 od 4 odprtih bralnih okvirjev kodirata za en protein zaradi puščajočega UAG stop kodona, njun produkt pa je replikaza z metilacijsko in helikazno podenoto, hkrati pa mala enota deluje kot zaviralec utišanja RNA, ki je obrambni mehanizem gostitelja. Movement protein (MP) pomaga pri sestavljanju novega viriona, v poznejših fazah replikacije, zapis zanj pa najdemo v subgenomski RNA. Prav tako za CP, kapsidni protein. | ||
Replikacija virusa poteka v citoplazmatski virusni tovarni (cytoplasmic viral factory) prek dvoverižnega intermediata. Ko plaščni proteini oddisociirajo se pozitivna virusna ssRNA začne prevajati na gostiteljskih ribosomih. Najprej se tako sintetizira virusna replikaza, ki potem sintetizira negativno verigo RNA. Ta je potem matrica za nadaljnjo sintezo pozitivne ssRNA, ki se vgradi v nove virione, ter subgenomske fragmente pozitivne RNA, iz katerih se izrazijo plaščni proteini in transportni proteini. Samo sestavljanje viriona poteka pod vplivom transportnih proteinov, ki enkapsidirajo ssRNA (+) v plaščne proteine. | Replikacija virusa poteka v citoplazmatski virusni tovarni (cytoplasmic viral factory) prek dvoverižnega intermediata. Ko plaščni proteini oddisociirajo se pozitivna virusna ssRNA začne prevajati na gostiteljskih ribosomih. Najprej se tako sintetizira virusna replikaza, ki potem sintetizira negativno verigo RNA. Ta je potem matrica za nadaljnjo sintezo pozitivne ssRNA, ki se vgradi v nove virione, ter subgenomske fragmente pozitivne RNA, iz katerih se izrazijo plaščni proteini in transportni proteini. Samo sestavljanje viriona poteka pod vplivom transportnih proteinov, ki enkapsidirajo ssRNA (+) v plaščne proteine. | ||
| Line 24: | Line 25: | ||
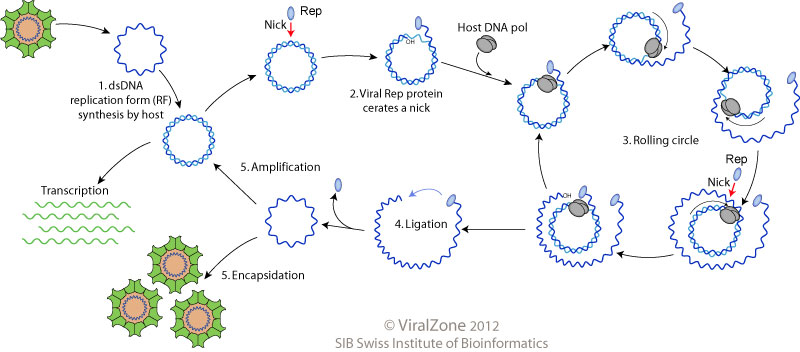
Replikacija virusne DNA poteka prek dvoverižnih intermediatov v jedru gostiteljske celice. Protein Rep ima ključno vlogo pri začetku in končanju replikacije; ustvari zarezo v krožno DNA, kjer potem gostiteljska polimeraza začne s transkripcijo, podvojeno DNA pa tudi poveže nazaj v krožno obliko. Samo podvojevanje poteka na način kotalečega se kroga. Ko virus vstopi v celico se kapsida odpre in v celico vstopi virusna ssDNA. Najprej se ta prek popravnih mehanizmov gostitelja pretvori v ds DNA (sinteza negativne verige). Potem, kot iniciator deluje Rep, ki ustvari zarezo v pozitivni verigi in gostiteljska polimeraza jo začne podvojevati. Po končanem ciklu Rep tudi ligira konce nove verige, in jo tako pripravi za enkapsidacijo v nove virione, cikel replikacije pa se ponavlja. Končni produkt je velika količina pozitivne ciklične ssDNA, ki se lahko enkapsidira v nove virione ali pa gre nazaj v replikacijski cikel (amplifikacija ali ojačanje). Mehanizem prehoda virusne DNA v sosednje celice pa še ni znan. | Replikacija virusne DNA poteka prek dvoverižnih intermediatov v jedru gostiteljske celice. Protein Rep ima ključno vlogo pri začetku in končanju replikacije; ustvari zarezo v krožno DNA, kjer potem gostiteljska polimeraza začne s transkripcijo, podvojeno DNA pa tudi poveže nazaj v krožno obliko. Samo podvojevanje poteka na način kotalečega se kroga. Ko virus vstopi v celico se kapsida odpre in v celico vstopi virusna ssDNA. Najprej se ta prek popravnih mehanizmov gostitelja pretvori v ds DNA (sinteza negativne verige). Potem, kot iniciator deluje Rep, ki ustvari zarezo v pozitivni verigi in gostiteljska polimeraza jo začne podvojevati. Po končanem ciklu Rep tudi ligira konce nove verige, in jo tako pripravi za enkapsidacijo v nove virione, cikel replikacije pa se ponavlja. Končni produkt je velika količina pozitivne ciklične ssDNA, ki se lahko enkapsidira v nove virione ali pa gre nazaj v replikacijski cikel (amplifikacija ali ojačanje). Mehanizem prehoda virusne DNA v sosednje celice pa še ni znan. | ||
[https://viralzone.expasy.org/resources/ssroll_replication.jpg Shema replikacije] po modelu kotalečega se kroga | |||
== Rastlinska plazmodezmata == | == Rastlinska plazmodezmata == | ||
Rastlinske celice so razvile način komunikacije na kratke in dolge razdalje za usklajeno regulacijo razvoja in prilagajanja na spremembe v okolju. Plazmodezmata (PD) so medcelični kanalčki, ki služijo za pretok majhnih molekul iz različnih bioloških procesov. Med razvojem rastlin je za razporeditev in diferenciacijo ključnega pomena transport majhnih RNA delcev (smRNA), transkripcijskih faktorjev in NCAP proteinov (''non-cell-autonomous protein''). Na prepustnost PD oz. SEL (''size exclusion limit'') vpliva količina kaloze (β-1,3` polisaharid glukoze) v celični steni, kjer njeno koncentracijo uravnavata kaloza sintaza in β-1,3`-glukanaza, ter nekateri proteini v PD. Tako lahko PD s spreminjanjem premera vpliva na difuzijo snovi preko celic. | Rastlinske celice so razvile način komunikacije na kratke in dolge razdalje za usklajeno regulacijo razvoja in prilagajanja na spremembe v okolju. [https://www.researchgate.net/profile/Shri_Yadav/publication/266624022/figure/fig1/AS:295628962844682@1447494845910/Plasmodesmata-Plasma-membrane-lined-pores-called-plasmodesmata-PD-penetrate-cell-walls.png Plazmodezmata] (PD) so medcelični kanalčki, ki služijo za pretok majhnih molekul iz različnih bioloških procesov. Med razvojem rastlin je za razporeditev in diferenciacijo ključnega pomena transport majhnih RNA delcev (smRNA), transkripcijskih faktorjev in NCAP proteinov (''non-cell-autonomous protein''). Na prepustnost PD oz. SEL (''size exclusion limit'') vpliva količina kaloze (β-1,3` polisaharid glukoze) v celični steni, kjer njeno koncentracijo uravnavata kaloza sintaza in β-1,3`-glukanaza, ter nekateri proteini v PD. Tako lahko PD s spreminjanjem premera vpliva na difuzijo snovi preko celic. | ||
Struktura PD je valjasto uvihana membrana, skozi katero poteka tanek tulec endoplazemskega retikuluma (dezmotubul) med obema celicama. Osnovna PD (''simple PD'') povezuje dve celici, medtem ko razvejana PD (''branched PD'') lahko povezuje več celic. PD lahko zavzame več konformacij, ki različno prepuščajo molekule oz. imajo različen SEL. Tako je omogočen selektiven transport majhnih molekul skozi PD in tudi večjih proteinov (do 10 kDa) skozi dezmotubul. Med razvojem je premer PD večji in lahko prepušča proteine tudi med 30 in 50 kDa, seveda pa je bolj od njihove mase pomembna oblika, saj se podolgovati proteini lažje spravijo skozi PD. | Struktura PD je valjasto uvihana membrana, skozi katero poteka tanek tulec endoplazemskega retikuluma (dezmotubul) med obema celicama. Osnovna PD (''simple PD'') povezuje dve celici, medtem ko razvejana PD (''branched PD'') lahko povezuje več celic. PD lahko zavzame več konformacij, ki različno prepuščajo molekule oz. imajo različen SEL. Tako je omogočen selektiven transport majhnih molekul skozi PD in tudi večjih proteinov (do 10 kDa) skozi dezmotubul. Med razvojem je premer PD večji in lahko prepušča proteine tudi med 30 in 50 kDa, seveda pa je bolj od njihove mase pomembna oblika, saj se podolgovati proteini lažje spravijo skozi PD. | ||
| Line 33: | Line 34: | ||
Širjenje virusa zaradi celične stene ni najbolj ugodno, zato so rastlinski virusi razvili mehanizme za prehajanje med celicami preko PD. Virusni gibalni proteini (movement protein-MP) imajo zmožnost povišanja SEL in s tem povečanja prepustnosti PD. Domnevajo, da na fronti širjenja virusa MP tvori kompleks s PD in s tem omogoči prost pretok ostalim molekulam virusa, a natančen opis mehanizma še ni znan. In vivo je znanih nekaj C-končnih fosforilacijskih mest na MP, ki bi lahko vplivala na regulacijo, kar podpira dejstvo, da je v PD povezanih več regulatornih kinaz. | Širjenje virusa zaradi celične stene ni najbolj ugodno, zato so rastlinski virusi razvili mehanizme za prehajanje med celicami preko PD. Virusni gibalni proteini (movement protein-MP) imajo zmožnost povišanja SEL in s tem povečanja prepustnosti PD. Domnevajo, da na fronti širjenja virusa MP tvori kompleks s PD in s tem omogoči prost pretok ostalim molekulam virusa, a natančen opis mehanizma še ni znan. In vivo je znanih nekaj C-končnih fosforilacijskih mest na MP, ki bi lahko vplivala na regulacijo, kar podpira dejstvo, da je v PD povezanih več regulatornih kinaz. | ||
Dodatna lastnost MP je zmožnost vezave enoverižne RNA neodvisno od zaporedja. Ker TMV (''tobacco mosaic virus'') za medcelično potovanje ne potrebuje plaščne ovojnice, se domneva, da v celici tvorita kompleks MP in virusna RNA (kompleks vRNP), kar omogoča lažje in hitrejše širjenje virusa po organizmu. Kompleks naj bi povzročil povišano delovanje glukanaze in s tem zmanjšanje kaloze | Dodatna lastnost MP je zmožnost vezave enoverižne RNA neodvisno od zaporedja. Ker TMV (''tobacco mosaic virus'') za medcelično potovanje ne potrebuje plaščne ovojnice, se domneva, da v celici tvorita kompleks MP in virusna RNA (kompleks vRNP), kar omogoča lažje in hitrejše širjenje virusa po organizmu. Kompleks naj bi povzročil povišano delovanje glukanaze in s tem zmanjšanje kaloze. | ||
Nekateri rastlinski virusi med celicami potujejo kot virioni, ki so večinoma večji od premera PD (>10nm). Taki virusi so razvili drugačen mehanizem prehajanja PD in sicer tako, da virusni MP tvorijo veliko tubularno strukturo, ki služi kot transport znotraj PD. V njih ni dezmotubula, tako da se lahko tubularna struktura umesti na sredino PD. Znano je, da C-terminalni deli MPjev interagirajo s plaščnimi proteini (''coat protein – CP'') znotraj tubula in tako omogočajo prehod viriona. Interakcija MP-CP je specifična za vsak virus posebej, kajti tubul iz MP enega virusa ne bo zmožen transporta nobenega virusa razen njemu lastnega. Iz teh povezav sklepajo, da sta formacija in transport analogna formaciji mikrotubulov (''thread milling''). | Nekateri rastlinski virusi med celicami potujejo kot virioni, ki so večinoma večji od premera PD (>10nm). Taki virusi so razvili drugačen mehanizem prehajanja PD in sicer tako, da virusni MP tvorijo veliko tubularno strukturo, ki služi kot transport znotraj PD. V njih ni dezmotubula, tako da se lahko tubularna struktura umesti na sredino PD. Znano je, da C-terminalni deli MPjev interagirajo s plaščnimi proteini (''coat protein – CP'') znotraj tubula in tako omogočajo prehod viriona. Interakcija MP-CP je specifična za vsak virus posebej, kajti tubul iz MP enega virusa ne bo zmožen transporta nobenega virusa razen njemu lastnega. Iz teh povezav sklepajo, da sta formacija in transport analogna formaciji mikrotubulov (''thread milling''). | ||
Latest revision as of 19:29, 5 May 2018
Rastlinski virusi se prenašajo med rastlinami preko rastlinskega soka, ko prideta v kontakt zdrava in poškodovana okužena rastlina (npr. pri prehranjevanju živali). Lahko pa se prenašajo tudi v insektnih vektorjih, kar jim omogoča prenos na daljše razdalje. Večina rastlinskih virusov ima za razliko od živalskih enoverižno RNA (ssRNA), ki je največkrat pozitivna (+ssRNA). Njihova oblika je večinoma paličasta s premerom do 20nm.
Modelni rastlinski virus
Tobacco mosaic virus
TMV je prvi odkriti rastlinski virus, ki pa se uporablja tudi kot modelni organizem. Zanimiv je tudi zaradi svojega promotorskega mesta, ki omogoča učinkovito ekspresijo proteinov v velikih količinah. TMV je zaradi tega dobra osnova za transformacijske vektorje za uporabo pri rastlinah. Tobacco mosaic virus iz rodu Tobamovirusov Tobamovirusi so edini rastlinski virusi paličaste oblike z enodelnim genomom. Virus se prenaša prek rastlinskih sokov in včasih tudi prek okuženih semen. Genom tega virusa obsega 4 odprte bralne okvirje in je dolg približno 6.0 do 6.5 kb v obliki linearne ssRNA. Na 5' koncu ima metilirano kapo (m7G5’pppG), 3' konec pa ima strukturo podobno tRNA. Zanimivo je to, da 2 od 4 odprtih bralnih okvirjev kodirata za en protein zaradi puščajočega UAG stop kodona, njun produkt pa je replikaza z metilacijsko in helikazno podenoto, hkrati pa mala enota deluje kot zaviralec utišanja RNA, ki je obrambni mehanizem gostitelja. Movement protein (MP) pomaga pri sestavljanju novega viriona, v poznejših fazah replikacije, zapis zanj pa najdemo v subgenomski RNA. Prav tako za CP, kapsidni protein.
Replikacija virusa poteka v citoplazmatski virusni tovarni (cytoplasmic viral factory) prek dvoverižnega intermediata. Ko plaščni proteini oddisociirajo se pozitivna virusna ssRNA začne prevajati na gostiteljskih ribosomih. Najprej se tako sintetizira virusna replikaza, ki potem sintetizira negativno verigo RNA. Ta je potem matrica za nadaljnjo sintezo pozitivne ssRNA, ki se vgradi v nove virione, ter subgenomske fragmente pozitivne RNA, iz katerih se izrazijo plaščni proteini in transportni proteini. Samo sestavljanje viriona poteka pod vplivom transportnih proteinov, ki enkapsidirajo ssRNA (+) v plaščne proteine.
Predstavnik ekonomsko najpomembnejših rastlinskih virusov
Rastlinski virusi pomembno vplivajo na svetovno proizvodnjo hrane in ekonomijo predvsem držav v razvoju, zato je njihovo razumevanje ključno pri zagotavljanju boljših življenjskih pogojev za ljudi po vsem svetu. Tomato yellow leaf curl virus je med bolj pomembnimi virusi glede na njegov negativen vpliv na pridelke paradižnika po svetu.
Tomato yellow leaf curl virus
Predstavnik skupine Begomovirusov iz družine Geminiviridae, ki se tako imenujejo zaradi svoje dvojne ikozaedrične zgradbe. Kapsida je sestavljena iz 22 pentamernih kapsomerov, ki so zgrajeni iz 110 kapsidnih proteinov. Prenaša se prek belih mušic Bemisia tabaci (whitefly), rastlinskega soka in semen. Obstaja več variacij virusa, glede na izvor, ki vse uporabljajo za vektor prenosa isto mušico, vendar pa imajo nekateri sevi dvodelni genom , nekateri pa enodelnega. TYLCV-TH (Tajski izolat) ima dve ciklični verigi ssDNA, ki sta skupaj dolgi približno 5.2 kb, TYLCV-IL (Izraelski sev) pa ima samo eno ciklično verigo ssDNA. Oba seva sta si med seboj podobna po proteomu, tu pa se bomo osredotočili na izraelski sev. Genom TYLCV-IL je sestavljen iz ene krožne pozitivne verige ssDNA dolge približno 2.7 kb, kodirne regije pa se nahajajo na obeh verigah, virionski (pozitivni) in negativni. Obsega 6 odprtih bralnih okvirjev , dva na virionski verigi (V1 in V2) in 4 na negativni verigi (C1, C2, C3 in C4), vsebuje pa tudi ohranjeno nukleotidno zaporedje, ki služi kot začetno mesto replikacije. V1 je kapsidni protein, ki ščiti virusno DNA, vloga V2, ki je nekakšen notranji ovoj, pa še ni znana. C1 je replikacijski protein, imenovan Rep, ki je edini protein potreben za replikacijo virusa. C2 protein deluje proti mehanizmom utišanja genov, s katerimi se rastline branijo proti virusnim okužbam, C3 protein je replikacijski ojačevalec in C4 determinanta nastopa simptomov okužbe.
Replikacija virusne DNA poteka prek dvoverižnih intermediatov v jedru gostiteljske celice. Protein Rep ima ključno vlogo pri začetku in končanju replikacije; ustvari zarezo v krožno DNA, kjer potem gostiteljska polimeraza začne s transkripcijo, podvojeno DNA pa tudi poveže nazaj v krožno obliko. Samo podvojevanje poteka na način kotalečega se kroga. Ko virus vstopi v celico se kapsida odpre in v celico vstopi virusna ssDNA. Najprej se ta prek popravnih mehanizmov gostitelja pretvori v ds DNA (sinteza negativne verige). Potem, kot iniciator deluje Rep, ki ustvari zarezo v pozitivni verigi in gostiteljska polimeraza jo začne podvojevati. Po končanem ciklu Rep tudi ligira konce nove verige, in jo tako pripravi za enkapsidacijo v nove virione, cikel replikacije pa se ponavlja. Končni produkt je velika količina pozitivne ciklične ssDNA, ki se lahko enkapsidira v nove virione ali pa gre nazaj v replikacijski cikel (amplifikacija ali ojačanje). Mehanizem prehoda virusne DNA v sosednje celice pa še ni znan.
Shema replikacije po modelu kotalečega se kroga
{kind=link}
Rastlinska plazmodezmata
Rastlinske celice so razvile način komunikacije na kratke in dolge razdalje za usklajeno regulacijo razvoja in prilagajanja na spremembe v okolju. Plazmodezmata (PD) so medcelični kanalčki, ki služijo za pretok majhnih molekul iz različnih bioloških procesov. Med razvojem rastlin je za razporeditev in diferenciacijo ključnega pomena transport majhnih RNA delcev (smRNA), transkripcijskih faktorjev in NCAP proteinov (non-cell-autonomous protein). Na prepustnost PD oz. SEL (size exclusion limit) vpliva količina kaloze (β-1,3` polisaharid glukoze) v celični steni, kjer njeno koncentracijo uravnavata kaloza sintaza in β-1,3`-glukanaza, ter nekateri proteini v PD. Tako lahko PD s spreminjanjem premera vpliva na difuzijo snovi preko celic. Struktura PD je valjasto uvihana membrana, skozi katero poteka tanek tulec endoplazemskega retikuluma (dezmotubul) med obema celicama. Osnovna PD (simple PD) povezuje dve celici, medtem ko razvejana PD (branched PD) lahko povezuje več celic. PD lahko zavzame več konformacij, ki različno prepuščajo molekule oz. imajo različen SEL. Tako je omogočen selektiven transport majhnih molekul skozi PD in tudi večjih proteinov (do 10 kDa) skozi dezmotubul. Med razvojem je premer PD večji in lahko prepušča proteine tudi med 30 in 50 kDa, seveda pa je bolj od njihove mase pomembna oblika, saj se podolgovati proteini lažje spravijo skozi PD.
{kind=link}
Premikanje virusov skozi PD
Širjenje virusa zaradi celične stene ni najbolj ugodno, zato so rastlinski virusi razvili mehanizme za prehajanje med celicami preko PD. Virusni gibalni proteini (movement protein-MP) imajo zmožnost povišanja SEL in s tem povečanja prepustnosti PD. Domnevajo, da na fronti širjenja virusa MP tvori kompleks s PD in s tem omogoči prost pretok ostalim molekulam virusa, a natančen opis mehanizma še ni znan. In vivo je znanih nekaj C-končnih fosforilacijskih mest na MP, ki bi lahko vplivala na regulacijo, kar podpira dejstvo, da je v PD povezanih več regulatornih kinaz. Dodatna lastnost MP je zmožnost vezave enoverižne RNA neodvisno od zaporedja. Ker TMV (tobacco mosaic virus) za medcelično potovanje ne potrebuje plaščne ovojnice, se domneva, da v celici tvorita kompleks MP in virusna RNA (kompleks vRNP), kar omogoča lažje in hitrejše širjenje virusa po organizmu. Kompleks naj bi povzročil povišano delovanje glukanaze in s tem zmanjšanje kaloze.
Nekateri rastlinski virusi med celicami potujejo kot virioni, ki so večinoma večji od premera PD (>10nm). Taki virusi so razvili drugačen mehanizem prehajanja PD in sicer tako, da virusni MP tvorijo veliko tubularno strukturo, ki služi kot transport znotraj PD. V njih ni dezmotubula, tako da se lahko tubularna struktura umesti na sredino PD. Znano je, da C-terminalni deli MPjev interagirajo s plaščnimi proteini (coat protein – CP) znotraj tubula in tako omogočajo prehod viriona. Interakcija MP-CP je specifična za vsak virus posebej, kajti tubul iz MP enega virusa ne bo zmožen transporta nobenega virusa razen njemu lastnega. Iz teh povezav sklepajo, da sta formacija in transport analogna formaciji mikrotubulov (thread milling).
Viri
[1] http://www.apsnet.org/edcenter/intropp/pathogengroups/pages/plantviruses.aspx (© 2018 The American Phytopathological Society. All rights reserved.) Introduction to Plant Viruses, the Invisible Foe Gergerich, R.C., and V. V. Dolja. 2006. Introduction to Plant Viruses, the Invisible Foe. The Plant Health Instructor. DOI: 10.1094/PHI-I-2006-0414-01
[2] https://viralzone.expasy.org/ (Swiss Institute of Bioinformatics) Viral Zone
[3] Rybicki, E. P. A Top Ten list for economically important plant viruses. Archives of Virology 160, 17–20 (2015).
[4] Heinlein, M. Plasmodesmata: Methods and protocols. Plasmodesmata: Methods and Protocols 1–346 (Springer New York, 2014). doi:10.1007/978-1-4939-1523-1