Mehanizem delovanja betalaktamov in glikopeptidov: Difference between revisions
Ajda Cafun (talk | contribs) |
Ajda Cafun (talk | contribs) |
||
| Line 15: | Line 15: | ||
== Mehanizem delovanja betalaktamskih antibiotikov == | == Mehanizem delovanja betalaktamskih antibiotikov == | ||
Betalaktamski antibiotiki, med katere spadajo penicilini, cefalosporini, | Betalaktamski antibiotiki, med katere spadajo penicilini, cefalosporini, karbapenemi (v množini pisani zato, ker obstaja več različnih modifikacij vsakega antibiotika) in drugi, so učinkovine, ki delujejo kot inhibitorji proteinov, ki vežejo penicilin. Potrebno je izpostaviti, da omenjene učinkovine ne delujejo na vse PBP, kar bi lahko sklepali iz imena teh proteinov, temveč na tiste, ki sodelujejo pri zadnjem koraku izgradnje celične stene, torej povezovanju peptidnih enot glikoproteinov. Z inhibicijo tega koraka vodijo celico v celično smrt. Delujejo tako na po Gramu pozitivne, kot tudi na po Gramu negativne bakterije. V večini primerov β-laktami inducirajo celično smrt prek razgradnje celične stene, obstajajo pa tudi primeri drugačnih mehanizmov celične smrti. Glavni tarčni protein β-laktamov, katerega inhibicija v največji meri pripomore k celični smrti, je DD-transpeptidaza, ki katalizira zadnji korak izgradnje celične stene, povezovanje dveh peptidnih verig. Encim ima v aktivnem mestu serinski ostanek, ki se kovalentno poveže s četrtim ostankom pentapeptida D-Ala, ob tem pa končni D-Ala izstopi iz peptidne verige. Na tako nastali acil-encim, se nato na končni D-Ala veže druga peptidna veriga preko L-lizinskega ali DAP ostanka na tretjem mestu. Povezani peptidni verigi izstopita iz aktivnega mesta encima. Tako je katalizirana povezava, ki zagotavlja čvrstost celične stene. | ||
Z razumevanjem mehanizma katalitične aktivnosti omenjenega PBP, lahko razumemo, zakaj so betalaktamski antibiotiki njihovi uspešni inhibitorji. Kot že ime pove, je ključna in skupna lastnost učinkovin β-laktamski obroč. Gre za štiričlenski obroč, ki vsebuje en dušikov atom, na enega od treh ogljikovih atomov pa je vezana karbonilna skupina. Pogosto je na β-laktamski obroč vezan še en obroč, kar poveča afiniteto njegove vezave v aktivno mesto encima. Penicilini na primer vsebujejo dodaten 5-členski obroč, cefalosporini pa 6-členski obroč. Oba dodatna obroča vsebujeta po en elektronegativen žveplov atom. Molekula β-laktama zaradi strukturne podobnosti s končnim delom pentapeptida D-Ala-D-Ala, ki je naravni substrat DD-peptidaz, posnema njegovo vezavo v aktivno mesto, pri čemer pa ima zaradi že omenjenih strukturnih lastnosti večjo afiniteto za vezavo. [[slika: http://tmedweb.tulane.edu/pharmwiki/lib/exe/fetch.php/betalactam_structure.png?w=600&tok=54cff1 ]] | Z razumevanjem mehanizma katalitične aktivnosti omenjenega PBP, lahko razumemo, zakaj so betalaktamski antibiotiki njihovi uspešni inhibitorji. Kot že ime pove, je ključna in skupna lastnost učinkovin β-laktamski obroč. Gre za štiričlenski obroč, ki vsebuje en dušikov atom, na enega od treh ogljikovih atomov pa je vezana karbonilna skupina. Pogosto je na β-laktamski obroč vezan še en obroč, kar poveča afiniteto njegove vezave v aktivno mesto encima. Penicilini na primer vsebujejo dodaten 5-členski obroč, cefalosporini pa 6-členski obroč. Oba dodatna obroča vsebujeta po en elektronegativen žveplov atom. Molekula β-laktama zaradi strukturne podobnosti s končnim delom pentapeptida D-Ala-D-Ala, ki je naravni substrat DD-peptidaz, posnema njegovo vezavo v aktivno mesto, pri čemer pa ima zaradi že omenjenih strukturnih lastnosti večjo afiniteto za vezavo. [[slika: http://tmedweb.tulane.edu/pharmwiki/lib/exe/fetch.php/betalactam_structure.png?w=600&tok=54cff1 ]] | ||
Latest revision as of 05:53, 15 May 2017
Sinteza bakterijske celične stene
Celična stena daje bakterijski celici trdnost in ščiti celično membrano pred poškodbami zaradi celičnega turgorja. V glavnem jo sestavlja polimer peptidoglikan, ki mu pravimo tudi murein. Po Gramu negativne bakterije imajo dve celični membrani, med katerima se v periplazemskem prostoru nahaja celična stena. Ta je široka od 7 nm do 8 nm in je sestavljena le iz nekaj plasti peptidoglikana. Po Gramu pozitivne bakterije nimajo dodatne zunanje celične membrane, celična stena, ki obdaja njihovo plazmalemo, pa je bolj debela. V širino meri od 20 nm do 80 nm, gradi pa jo lahko do 40 plasti peptidoglikana.
Peptidoglikan je zgrajen iz vzporednih sladkornih verig, povezanih preko oligopeptidov. V sladkornem ogrodju so izmenjujoče nanizani ostanki N-acetilglukozamina (NAG) in N-acetilmuraminske kisline (NAM), ki so povezani z β(1 → 4) glikozidno vezjo. Na D-laktilno skupino vsakega ostanka NAM je vezan tetrapeptid. Aminokisline v tem peptidu si sledijo po zaporedju: L-alanin, D-glutamin, 2,6-diaminopimelinska kislina (DAP) in D-alanin pri po Gramu negativnih bakterijah, pri po Gramu pozitivnih bakterijah pa je na tretjem mestu namesto DAP L-lizin. Pri po Gramu pozitivnih bakterijah ta povezava ni direktna, ampak poteka preko peptidnih mostičkov, ki so ponavadi sestavljeni iz petih zaporednih glicinskih ostankov.
Pri sintezi celične stene se sestavni elementi peptidoglikana izgradijo v citosolu, nato pa se prenesejo v periplazmo, kjer se povežejo med seboj. Na začetku se na molekulo NAM postopoma pripne 5 aminokislinskih ostankov. Prvi štirje si sledijo po prej navedenem zaporedju, kot peti pa je pripet še en dodaten D-alanin. Nastala molekula se nato preko NAM pripne na lipid C55-izoprenil pirofosfat, imenovan tudi baktrofenil, ki se nahaja v proti citosolu usmerjeni polovici celične membrane. Z NAM se v naslednjem koraku poveže NAG, na peptidni del pa se lahko doda še povezovalni glicinski mostiček. Celotna molekula se sedaj iz notranje polovice lipidnega dvosloja ob pomoči flipaze FtsW prestavi v zunanjo polovico, tako da sladkorno-peptidni del gleda v periplazemski prostor. V periplazmi poteka povezovanje peptidoglikanskih enot s pomočjo transglikozilaz in transpeptidaz, ki jih z nadpomenko imenujemo penicilin-vezavni proteini (PBP). Transglikozilaza prepozna na lipidno komponento vezan peptidoglikanski prekurzor in poveže četrti C-atom ostanka NAG na prekurzorju s prvim C-atomom NAM, ki se nahaja na koncu izgrajujoče peptidoglikanske verige. Pri tem nastane β(1 → 4) vez, baktrofenil pa se odcepi in prenese nazaj v notranjo polovico fosfolipidnega dvosloja, kjer je pripravljen na sprejem novega prekurzorja. Sedaj se morajo oligopeptidi še prečno povezati, kar katalizira transpeptidaza. Encim odcepi končni D-alanin s pentapeptida na prekurzorju in D-alanin na preostalem tetrapeptidu poveže s tretjim aminokislinskim ostankom v sosednjem tetrapeptidu, bodisi direktno, bodisi preko vmesnega glicinskega mostička.
Bakterije lahko lastno celično steno tudi razgrajujejo. Pri tem sodelujejo avtolizini, ki so potrebni za obnavljanje in spreminjanje oblike celične stene, ter regulacijo njene rasti. Procesa izgrajevanja in razgrajevanja sta uravnotežena, ob določenih pogojih pa avtolizini sprožijo intenzivnejšo razgradnjo stene, ki vodi v celično lizo. Ta je lahko tudi posledica delovanja antibiotikov.
Mehanizem delovanja betalaktamskih antibiotikov
Betalaktamski antibiotiki, med katere spadajo penicilini, cefalosporini, karbapenemi (v množini pisani zato, ker obstaja več različnih modifikacij vsakega antibiotika) in drugi, so učinkovine, ki delujejo kot inhibitorji proteinov, ki vežejo penicilin. Potrebno je izpostaviti, da omenjene učinkovine ne delujejo na vse PBP, kar bi lahko sklepali iz imena teh proteinov, temveč na tiste, ki sodelujejo pri zadnjem koraku izgradnje celične stene, torej povezovanju peptidnih enot glikoproteinov. Z inhibicijo tega koraka vodijo celico v celično smrt. Delujejo tako na po Gramu pozitivne, kot tudi na po Gramu negativne bakterije. V večini primerov β-laktami inducirajo celično smrt prek razgradnje celične stene, obstajajo pa tudi primeri drugačnih mehanizmov celične smrti. Glavni tarčni protein β-laktamov, katerega inhibicija v največji meri pripomore k celični smrti, je DD-transpeptidaza, ki katalizira zadnji korak izgradnje celične stene, povezovanje dveh peptidnih verig. Encim ima v aktivnem mestu serinski ostanek, ki se kovalentno poveže s četrtim ostankom pentapeptida D-Ala, ob tem pa končni D-Ala izstopi iz peptidne verige. Na tako nastali acil-encim, se nato na končni D-Ala veže druga peptidna veriga preko L-lizinskega ali DAP ostanka na tretjem mestu. Povezani peptidni verigi izstopita iz aktivnega mesta encima. Tako je katalizirana povezava, ki zagotavlja čvrstost celične stene.
Z razumevanjem mehanizma katalitične aktivnosti omenjenega PBP, lahko razumemo, zakaj so betalaktamski antibiotiki njihovi uspešni inhibitorji. Kot že ime pove, je ključna in skupna lastnost učinkovin β-laktamski obroč. Gre za štiričlenski obroč, ki vsebuje en dušikov atom, na enega od treh ogljikovih atomov pa je vezana karbonilna skupina. Pogosto je na β-laktamski obroč vezan še en obroč, kar poveča afiniteto njegove vezave v aktivno mesto encima. Penicilini na primer vsebujejo dodaten 5-členski obroč, cefalosporini pa 6-členski obroč. Oba dodatna obroča vsebujeta po en elektronegativen žveplov atom. Molekula β-laktama zaradi strukturne podobnosti s končnim delom pentapeptida D-Ala-D-Ala, ki je naravni substrat DD-peptidaz, posnema njegovo vezavo v aktivno mesto, pri čemer pa ima zaradi že omenjenih strukturnih lastnosti večjo afiniteto za vezavo. [[slika: http://tmedweb.tulane.edu/pharmwiki/lib/exe/fetch.php/betalactam_structure.png?w=600&tok=54cff1 ]] Za razliko od naravnega substrat, ki z vezavo na aktivni serin omogoča izstop končnega ostanka iz pentapeptida, pa po vezavi β-laktamskega obroča ni možen izstop dela molekule, saj je elektrofilni C-atom, na katerega se -OH skupina serina veže, del obroča. Molekula ostane tako kovalentno vezana na aktivno mesto, ki s tem ni več zmožno opravljati svoje katalitične funkcije.
Različni betalaktamski antibiotiki imajo različne afinitete do enega ali več PBP, zato sprožijo različne morfološke spremembe bakterije. Proteini, ki spadajo v skupino PBP1, med drugim DD-transpeptidaza, vplivajo na elongacijo celice in njihova inhibicija v največji meri pripomore k lizi celične stene. Inhibicija PBP3 proteinov, ki vplivajo na celično delitev, povzroči filamentacijo celice, kar pomeni abnormalno elongacijo, pri čemer se celica ne deli. Inhibicija proteinov PBP3 je povezana z lizo, vendar v manjši meri kot inhibicija proteinov PBP1. Inhibicija proteinov PBP2, ki skrbijo za obliko celice, ne vodi v lizo celične stene. Ker je za lizo poleg inhibicije izgradnje celične stene potrebna tudi njena razgradnja s strani avtolizinov, so znanstveniki potrdili, da delovanje inhibitorjev proteinov PBP1 v največji meri spodbudi delovanje avtolizinov, delovanje inhibitorjev proteinov PBP2 pa v najmanjši meri.
Mehanizem delovanje glikopeptidnih antibiotikov
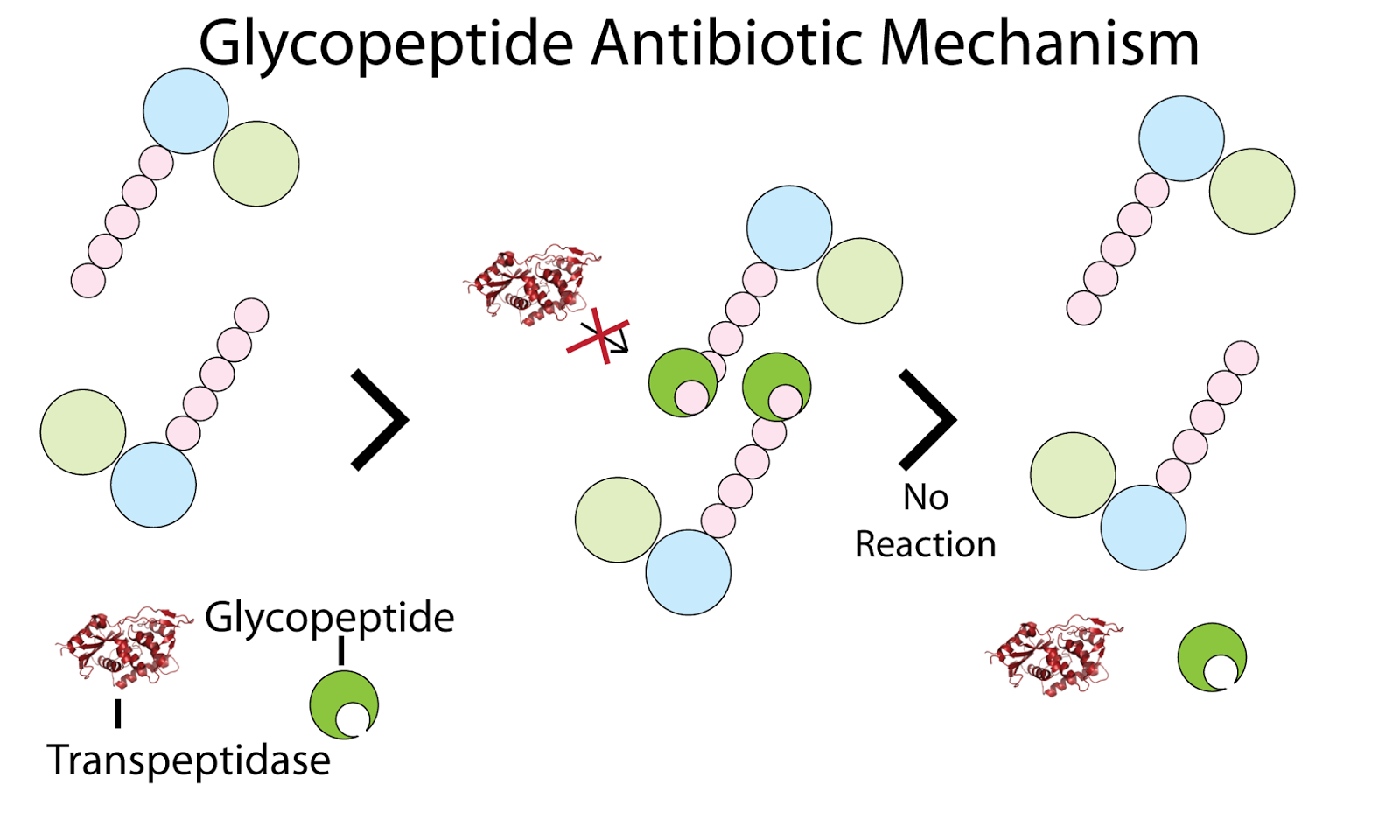
Glikopeptidi so glikozilirani neribosomski peptidi, ki delujejo kot sterični inhibitorji zorenja peptidoglikanov grampozitivnih bakterij. Inhibirajo lahko delovanje dveh encimov, in sicer transglikozilaze in transpeptidaze. Ta inhibicija ne poteka preko vezave na encim, pač pa se glikopeptidi namesto tega vežejo na substrat omenjenih encimov, peptidoglikan, natančneje na njihov oligopeptidni del. Vezani glikopeptidi predstavljajo sterično oviro in encimoma preprečujejo dostop do peptidoglikana. Na ta način preprečujejo ustrezno povezovanje peptidoglikana. Posledično je inhibirana rast, smrt pa po navadi povzroči povečan tlak, ki nastane zaradi nezadostno povezanega peptidoglikana. Pride do osmotske lize. [[slika: http://4.bp.blogspot.com/-aMRnL9j_6ZY/UfVm0sjiUYI/AAAAAAAAA50/nvP133cuItU/s1600/transpeptidase-07.png ]]
Inhibicija preko vezave na substrat je mogoča zaradi posebne strukture glikopeptidov. Sestavljeni so iz osrednjega heptapeptida, ki ga tvorijo pretežno aromatske aminokisline. Na heptapeptidno ogrodje so vezane različne skupine, najpogosteje sladkorni ostanki, kloridni atomi in lipidne verige. Število aromatskih aminokislin v heptapetidu se med posameznimi glikopeptidi lahko razlikuje. V precej poznanem peptidnem antibiotiku, vankomicinu, je aromatskih aminokslin 5, medtem ko jih je v teikoplaninu 7. Aromatske aminokisline v osrednjem peptidu so oksidativno prečno povezane. Prečne povezave tvorijo posebno strukturno konformacijo, »čeljust«, v katero se vežejo peptidni deli peptidoglikana. [[slika: https://www.researchgate.net/figure/50893040_fig6_Figure-1-Chemical-structures-of-Type-I-1-vancomycin-2-balhimycin-and-Type-IV-3 ]]
Glikopeptidi se s čeljustmi vežejo na C-konce peptidoglikana, ki so ključni za tvorbo kompleksa transglikozilaza-peptidoglikan ali transpeptidaza-peptidoglikan. V splošnem glikopeptidi prepoznajo skupine aminokislin s stereokemijsko konformacijo -L-D-D, najmanjša enota, na katero se lahko vežejo, pa je -D-ala-D-ala. Za C-konce nastajajoče konce peptidoglikana, je značilno zaporedje L-lys-D-ala-D-ala, preko katerega transglikozilaza poveže dve peptidoglikanski enoti, transpeptidaza pa preko njega nove peptide vgrajuje v celično steno. Glikopeptidi z zaporedjem –D-ala-D-ala v peptidnem delu peptidoglikana tvorijo vodikove vezi in se na ta način, kljub svoji siceršnji rigidnosti, ovijejo okoli peptidoglikana, kar preprečuje vezavo encimov. Vodikove vezi po navadi nastanejo med C-končno karboksilno skupino motiva -D-ala-D-ala in amidnimi skupinami aminokislin iz heptapeptidnega ogrodja glikopeptidov. Mogoč je tudi nastanek vodikove vezi med aminsko skupino končnega alanina v motivu –D-ala-D-ala in kisikom karbonilne skupine osrednjega peptida. [[slika: http://offset.peczuh.uconn.edu/wp-content/uploads/sites/240/2015/08/b100912p-f2.gif. ]]
Večina glikopeptidnih antibiotikov lahko preko tvorbe vodikovih vezi tudi homodimerizira. Takšna homodimerizacija je skoraj vedno, izjema je le rizocitin, kooperativna z vezavo na zaporedje -D-ala-D-ala. Kooperativnost naj bi bila posledica tega, da vezava peptidoglikana v strukturo glikopeptida uvede simetrijo, ki omogoči tvorbo več intramolekularnih vezi. Slednje spremenijo konformacijo glikopeptida tako, da tesneje ovije peptid in tvori nove vezi. Dimerizacija predstavlja tudi večjo sterično oviro za transpeptidazo in transglikozilazo. Glikopeptidni antibiotiki, ki imajo večjo tandenco za dimerizacijo, so načeloma učinkovitejši. Teikoplanin je najbolj znan antibiotik iz družine glikopeptidov, ki ne tvori homodimerov. Kljub temu je njegovo delovanje zelo učinkovito, saj vsebuje dodatno vezano N-subsituirano maščobnokislinsko stransko verigo. Ta je vezana preko glukozaminskega ostanka, ki je vezan na 4. aminokislino osrednjega heptapeptida. Iz tega je jasno razvidno, da na učinkovitost antibiotika vplivajo tudi njegove stranske skupine. Hidrofobne stranske verige v splošnem omogočajo boljšo lokalizacijo in sidranje antibiotika na tarčno mesto, še pomembneje pa je, da omogočajo precej močne intramolekularne interakcije, katerih vpliv je pojasnjen nekoliko višje v besedilu. [[slika: http://www.hull.ac.uk/php/chsanb/LMWeb/Image84.gif ]] Rezistentne bakterije se učinku gikopeptidov lahko izognejo tako, da namesto zadnjega D-alanina v oligopeptide vgrajujejo D-laktat. Na takšne oligopeptide se glikopeptidi ne vežejo.
Viri
- Kang H-K, Park Y. Glycopeptide Antibiotics: Structure and Mechanisms of Action. J Bacteriol Virol. 2015;45(2):67-78. http://synapse.koreamed.org/DOIx.php?id=10.4167%2Fjbv.2015.45.2.67.
- Höltje, J. V. Growth of the stress-bearing and shape-maintaining murein sacculus of Escherichia coli. Microbiol. Mol. Biol. Rev. 62, 181–203 (1998).

{kind=link}
{kind=link}
{kind=link}
{kind=link}