Celični skelet
Glavne naloge citoskeleta
- Zgradba in opora
- Znotraj celični transport
- Krčljivost in gibanje
- Prostorska ureditev
{kind=link}
Aktinski filamenti
Aktinski filamenti so eni od treh tipov citoskeleta. Zgrajeni so iz dveh prepletajočih se verig in najdemo jih pod celično membrano. Polimerno obliko aktina imenujemo F-aktin (d = 7 mm), monomerno pa G-aktin.
Vloga aktina v celici:
{kind=link}
- mišična kontrakcija
- ameboidno gibanje
- celična delitev
- celična lokomocija
- vzdrževanje oblike celice
Aktin je sestavljen iz 375 aminokislin in je najbolj zastopan protein v celici.
Poznamo 6 izomernih oblik:
- 4-α-aktin: v celicah za krčenje
- 1-β-aktin: v celicah za gibanje
- 1-γ-aktin: v gladkih mišičnih celicah
- Arp 1 (actin related protein): vključen je v transport povezan z mikrotubuli
- Arp 2/3: se povežeta v kompleks; sodelujeta pri nastajanju in razvejanju aktinskih filamentov
Nastanek aktinskih filamentov:
Odvisen od zadostne koncentracije G-aktina z vezanim ATP ter ustreznega ionskega razmerja.
Aktinska molekula je globularni protein (G-aktin). V svoji zgradbi vsebuje posebno mesto za vezavo ATP ali ADP. Vezava je odvisna od koncentracije Mg2+ ionov. Po vezavi ATP ima G-aktin 10-krat večjo afiniteto do vezave z drugo molekulo G-aktina. Na ta način nastane iz monomera polimer (F-aktin). Koncu, kjer se molekule dodajajo, pravimo pozitivni konec. Na nasprotnem, negativnem koncu pa prihaja do hidrolize ATP v ADP. Molekule z vezanim ADP pa se hitreje odcepljajo. Prenos monomera s + konca na – konec na F-aktinu imenujemo 'treadmilling'. V primeru povišane koncentracije G-aktina (> 0,8 μM) poteka dodajanje na obeh koncih filamenta. Če pa je koncentracija manjša od 0,1 μM, se začne filament krajšati na obeh koncih.
Pri vzdrževanju razmerja med monomerno in polimerno obliko sodelujeta 2 proteina: TIMOZIN in PROFILIN, pri cepitvi pa KOFILIN in GELSOLIN. Proteine, ki stabilizirajo aktinske filamente, imenujemo pokrovni ali capping proteini. Najbolj znana sta: CAP Z in TROPOMODULIN.
{kind=link}
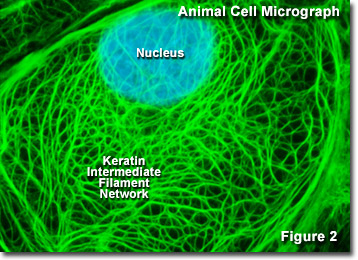
Intermediarni filamenti
Intermediarni filamenti so heterogena skupina citoskeleta (poznamo že okoli 70 genov), ki so celično specifični. So dolgi, nitasti filamenti s premerom 10 nm, ki izhajajo iz centra celice proti membrani. Ne potrebujejo ne ATP-ja ne GTP-ja za svoj obstoj. Filamenti niso polarizirani in so odporni na razteznostne sile, zato so zelo pomembni v epitelnem in mišičnem tkivu.
Njihova vloga:
{kind=link}
- celična in tkivna integriteta
- lokalizacija in delovanje celičnih organelov:
- vzdrževanje mitohondrijev ob miofibrilah v mišični celici
- vzdrževanje Golgijevega aparata ob celičnem jedru
- razporejanje lizosomov
- povezovanje jedra s transmembranskimi proteini
- vzdrževanje oblike jedra (jedrna lamina)
- usmerjanje proteinov na tarčna mesta
- vzdrževanje mitohondrijev ob miofibrilah v mišični celici
Delimo jih na 6 skupin:
- kisli keratini
- bazični keratini
- vimentin (krvne celice in fibroblasti), dezmin (mišične celice), gliakisli proteini in periferin (živčne celice)
- lahki NF-L, srednje težki NF-M, težki NF-H: pomembni za vzdrževanje aksonov
- lamini A,B,C: sestavljajo jedrno lamino, ki predstavlja bazo, na kateri se gradi jedrna membrana. Lamina A in C se povezujeta v mrežo, lamin B pa predstavlja povezavo z jedrno membrano
- nestin: pomemben med embrionalnim razvojem (nastajanje nevronov v matičnih celicah)
Intermediarni filamenti so sestavljeni iz treh regij:
Glava, ki je na N-koncu; osrednja paličasta regija, ki predstavlja konzervativni del proteina; in rep, ki je na C-koncu.
Pomemben povezovalni protein je plektin, ki povezuje intermediarne filamente z drugimi elementi citoskeleta.
{kind=link}
Mikrotubuli
Mikrotubuli so dolge cevaste strukture s premerom 24 nm. Sestavljeni so iz tubulinskih dimerov α in β. Ti se povežejo v linearne protofilamente. 13 paralelno v krog vezanih protofilamentov tvori en mikrotubul. Mikrotubuli so polarni - na β-koncu pozitivni in negativni na α-koncu. Rast mikrotubulov se začne na z γ-tubulini obdanem centrosomu. Negativni α-konec se prvi veže, zato mikrotubuli rastejo proti pozitivnemu koncu. Za svojo rast potrebujejo molekule GTP. Na α-mesto se veže GTP, da poveže obe enoti v dimer. Na β-mesto pa se veže GTP, ki trdno povezuje druge dimere. Vendar se lahko na β-mestu GTP hidrolizira v GDP in oslabi vez. Hitrost rasti je odvisna od koncentracije tubulinskih dimerov. Če se hidroliza GTP v GDP nadaljuje vse do konca mikrotubula, se zgodi hitra razgradnja na tubulinske dimere. Temu pojavu pravimo dinamična nestabilnost.
Vloga mikrotubulov:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
- celična in tkivna integriteta
- zagotavljajo znotrajcelično vezavo za motorne proteine KINEZINE in DINEINE
- delitev kromosomov pri mitozi
- oblikovanje delitvenega vretena
- sestavljajo bazalno telo (migetalke) - z gibanjem premikajo tekočino
- sestavljajo aksonemo bičkov (1-2 na celico), ki utripajo v valovih in prenašajo tekočino
{kind=link}
{kind=link}
Motorni proteini:
Kinezini se premikajo proti pozitivnemu koncu, dineini pa proti negativnemu koncu, oba proteina sodelujeta pri prenašanju organelov, delitvi celice in premikanju bičkov ali migetalk.
Proteini, ki sodelujeo pri rasti mikrotubula:
XMAP 215 se vežajo na + konec in ga stabilizirajo. KATASTROFIN hitro odceplja kompomente. KATANIN cepi mikrotubul po celi dolžini.
{kind=link}