Metilacijski sistem pri bakterijah: Difference between revisions
Maks Kumek (talk | contribs) |
Maks Kumek (talk | contribs) |
||
| (6 intermediate revisions by 2 users not shown) | |||
| Line 1: | Line 1: | ||
== Uvod == | == Uvod == | ||
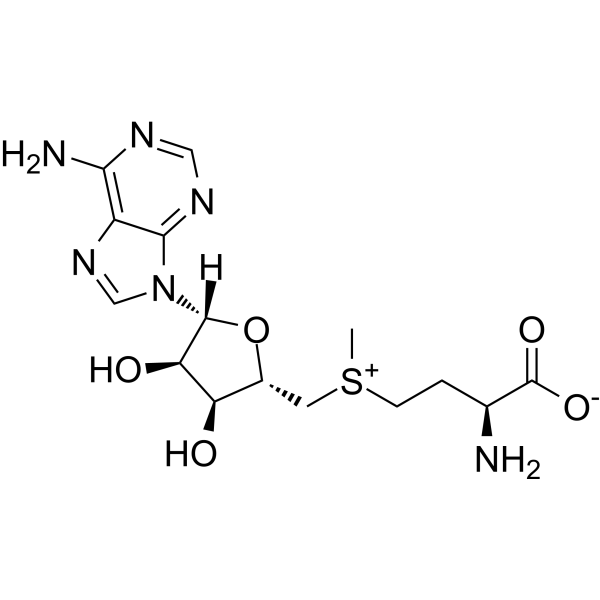
Metilacija DNA je eden najpomembnejših načinov modifikacije DNA in je prisotna v genomih različnih organizmov. Metilacija DNA pri bakterijah vpliva na različne | Metilacija DNA je eden izmed najpomembnejših načinov modifikacije DNA in je prisotna v genomih različnih organizmov. Metilacija DNA pri bakterijah vpliva na različne procese, kot so: genska regulacija, kromosomsko podvojevanje/popravljanje, prepoznavanje tuje DNA v gostiteljski celici in posledično rezanje tuje DNA ter v posebnem primeru preživetje virusne DNA v gostiteljski celici. Pri bakterijah do metilacije DNA prihaja na bazah v nukleotidih adenina in citozina. Metiltransferaze (MTaz) katalizirajo ta postopek, pri čemer je S-adenozil-L-metionin ([https://file.medchemexpress.com/product_pic/hy-b0617.gif AdoMet]) univerzalni donor metilne skupine. Po reakciji nastane metilirana DNA in AdoHcy (S-adendozil-L-homocistein) [1]. | ||
Na osnovi znanih funkcij bakterijskih metiltransferaz, jih delimo v tri velike skupine: | Na osnovi znanih funkcij bakterijskih metiltransferaz, jih delimo v tri velike skupine: | ||
• MTaze, ki niso del restrikcijo- | • MTaze, ki niso del restrikcijo-modifikacijskih sistemov, vendar lahko sodelujejo pri celični regulaciji, | ||
• MTaze, ki spadajo v restrikcijsko- | • MTaze, ki spadajo v restrikcijsko-modifikacijske sisteme, | ||
• virusne MTaze, ki jih prenašajo fagi in jih | • virusne MTaze, ki jih prenašajo fagi in jih uporabljajo za povečanje obstoja svoje DNA v gostiteljskih celicah. | ||
Bakterije imajo ti. restrikcijsko-modifikacijske sisteme. DNA MTaze imajo v njih vlogo zaščite lastne DNA pred restrikcijskimi encimi. R/M sistemi delujejo namreč tako, da metilirajo na določenem zaporedju DNA, ki ga prepoznavajo tudi restrikcijski encimi. Preprečena je vezava restrikaz in cepljenje lastne dvojne vijačnice. S tem bakterija prepreči zase usoden dogodek [1]. | Bakterije imajo ti. restrikcijsko-modifikacijske sisteme. DNA MTaze imajo v njih vlogo zaščite lastne DNA pred restrikcijskimi encimi. R/M sistemi delujejo namreč tako, da metilirajo na določenem zaporedju DNA, ki ga prepoznavajo tudi restrikcijski encimi. Preprečena je vezava restrikaz in cepljenje lastne dvojne vijačnice. S tem bakterija prepreči zase usoden dogodek [1]. | ||
| Line 17: | Line 17: | ||
DNA adenin MTaza (Dam) je eden izmed encimov velike skupine metiltransferaz, ki niso del R/M sistema. Dam metilira na adeninu in igra vlogo pri metiliranju hemimetilirane DNA, ki nastane pri replikaciji. | DNA adenin MTaza (Dam) je eden izmed encimov velike skupine metiltransferaz, ki niso del R/M sistema. Dam metilira na adeninu in igra vlogo pri metiliranju hemimetilirane DNA, ki nastane pri replikaciji. | ||
Na hemimetilirano DNA se na promotorski regiji vežejo DNA vezavni proteini SeqA. Ko je SeqA vezan, preprečuje vezavo Dam na promotorsko regijo in posledično metilacijo samega promotorja. Slednje ima posledico inhibicije ali indukcije transkripcije genov za različne proteine. Spontana disocijacija SeqA dovoli dostop in vezavo Dam na DNA. Ko veže promotor, začne drseti po vijačnici dokler ne pride do specifičnega nukleotidnega zaporedja 5' – GATC – 3' in metilira adenin. Stabilnost adenina v aktivnem mestu omogoči DPPY motiv na proteinu. Po [http://mol-biol4masters.masters.grkraj.org/html/Prokaryotic_DNA_Replication3-E_coli_DNA_Replication_files/image003.jpg metiliranju promotorja] se lahko veže bakterijska dnaA, ki je transkripcijski aktivator. | Na hemimetilirano DNA se na promotorski regiji vežejo DNA vezavni proteini SeqA. Ko je SeqA vezan, preprečuje vezavo Dam na promotorsko regijo in posledično metilacijo samega promotorja. Slednje ima posledico inhibicije ali indukcije transkripcije genov za različne proteine. Spontana disocijacija SeqA dovoli dostop in vezavo Dam na DNA. Ko veže promotor, začne drseti po vijačnici dokler ne pride do specifičnega nukleotidnega zaporedja 5' – GATC – 3' in metilira adenin. Stabilnost adenina v aktivnem mestu omogoči DPPY motiv na proteinu. Po [http://mol-biol4masters.masters.grkraj.org/html/Prokaryotic_DNA_Replication3-E_coli_DNA_Replication_files/image003.jpg metiliranju promotorja] se lahko veže bakterijska dnaA, ki je transkripcijski aktivator [2]. | ||
== Zgradba metiltransferaz == | == Zgradba metiltransferaz == | ||
| Line 30: | Line 29: | ||
TRD domena prepozna, stabilizira in veže DNA, vendar njeno aminokislinsko zaporedje ni ohranjeno. Pač pa je eno najbolj diverznih zaporedji aminokislinskih ostankov v encimu. Velikokrat so na robu teh zaporedji ponavljajoča se dokaj ohranjena zaporedja. Pomembno je poudariti, da ni direktne povezave med enim aminokislinskim ostankom in enim nukleotidom, marveč so interakcije med TRD in DNA kompleksne narave. | TRD domena prepozna, stabilizira in veže DNA, vendar njeno aminokislinsko zaporedje ni ohranjeno. Pač pa je eno najbolj diverznih zaporedji aminokislinskih ostankov v encimu. Velikokrat so na robu teh zaporedji ponavljajoča se dokaj ohranjena zaporedja. Pomembno je poudariti, da ni direktne povezave med enim aminokislinskim ostankom in enim nukleotidom, marveč so interakcije med TRD in DNA kompleksne narave. | ||
AdoMet vezavna domena veže na kofaktor. Sestavljena je iz dveh poddomen: ena za vezavo AdoMet in druga za vezavo na prejemnika metilne skupine. AdoMet ima najpomembnejši motiv I. Na osnovi aktivnosti AdoMet vezavne in katalitične domene delimo metiltransferaze R/M | AdoMet vezavna domena se veže na kofaktor. Sestavljena je iz dveh poddomen: ena za vezavo AdoMet in druga za vezavo na prejemnika metilne skupine. AdoMet ima najpomembnejši motiv I. Na osnovi aktivnosti AdoMet vezavne in katalitične domene delimo metiltransferaze R/M sistemov na: eksociklične amino MTaze (N6-adenin MTaza in N4-citozin MTaza) in endociklične MTaze (C5-citozin MTaze) [3]. | ||
== C5 metilacija == | == C5 metilacija == | ||
| Line 50: | Line 49: | ||
== Obračanje baz == | == Obračanje baz == | ||
Pri vseh metiltransferazah | Pri vseh metiltransferazah je potreba po premestitvi baze, ki se metilira, velika. Razlogi so predvsem sterične narave. Pri C5 metilaciji npr. je to nastanek intermediatov, ki nimajo prostora v klasični B-obliki DNA vijačnice. Tako je bila že prav pri modelu kompleksa HhaI z AdoMet odkrita oblika, kjer je baza, ki je namenjena metilaciji, rotirana iz osi dvojne vijačnice za 180°. Ta baza je tako segala iz malega žleba DNA v katalitično domeno encima. Ista potreba po rotaciji baz se je pokazala pri vseh ostalih vrstah metilaz. | ||
Osrednjo vlogo v sprostitvi baze iz DNA ima TRD. [https://media.nature.com/m685/nature-assets/nsmb/journal/v8/n2/images/nsb0201_121_F1.jpg M.TaqI] modeli so pokazali, da se ob vezavi struktura vijačnice močno deformira. Razmakne se mali žleb za približno 3 °A, stisne se ogrodje in | Osrednjo vlogo v sprostitvi baze iz DNA ima TRD. [https://media.nature.com/m685/nature-assets/nsmb/journal/v8/n2/images/nsb0201_121_F1.jpg M.TaqI] modeli so pokazali, da se ob vezavi struktura vijačnice močno deformira. Razmakne se mali žleb za približno 3 °A, stisne se ogrodje in timidin se pomakne bolj proti centru vijačnice. To je posledica interakcij med skoraj vsemi baznimi pari [https://media.nature.com/m685/nature-assets/nsmb/journal/v8/n2/images/nsb0201_121_F2.gif prepoznavnega zaporedja DNA], razen timidina, ki je par metiliranemu adenozinu. Vse to pripomore prekinitvi van der Waalsovih vezi med tem parom in izpostavitvi adenozina [4]. | ||
== Metiltransferaze restrikcijsko-modifikacijskih sistemov (R/M sistemi) == | == Metiltransferaze restrikcijsko-modifikacijskih sistemov (R/M sistemi) == | ||
Latest revision as of 19:14, 9 April 2019
Uvod
Metilacija DNA je eden izmed najpomembnejših načinov modifikacije DNA in je prisotna v genomih različnih organizmov. Metilacija DNA pri bakterijah vpliva na različne procese, kot so: genska regulacija, kromosomsko podvojevanje/popravljanje, prepoznavanje tuje DNA v gostiteljski celici in posledično rezanje tuje DNA ter v posebnem primeru preživetje virusne DNA v gostiteljski celici. Pri bakterijah do metilacije DNA prihaja na bazah v nukleotidih adenina in citozina. Metiltransferaze (MTaz) katalizirajo ta postopek, pri čemer je S-adenozil-L-metionin (AdoMet) univerzalni donor metilne skupine. Po reakciji nastane metilirana DNA in AdoHcy (S-adendozil-L-homocistein) [1].
Na osnovi znanih funkcij bakterijskih metiltransferaz, jih delimo v tri velike skupine:
• MTaze, ki niso del restrikcijo-modifikacijskih sistemov, vendar lahko sodelujejo pri celični regulaciji,
• MTaze, ki spadajo v restrikcijsko-modifikacijske sisteme,
• virusne MTaze, ki jih prenašajo fagi in jih uporabljajo za povečanje obstoja svoje DNA v gostiteljskih celicah.
Bakterije imajo ti. restrikcijsko-modifikacijske sisteme. DNA MTaze imajo v njih vlogo zaščite lastne DNA pred restrikcijskimi encimi. R/M sistemi delujejo namreč tako, da metilirajo na določenem zaporedju DNA, ki ga prepoznavajo tudi restrikcijski encimi. Preprečena je vezava restrikaz in cepljenje lastne dvojne vijačnice. S tem bakterija prepreči zase usoden dogodek [1].
Delovanje metiltransferaz, ki niso del restrikcijsko-metilacijskega sistema
DNA adenin MTaza (Dam) je eden izmed encimov velike skupine metiltransferaz, ki niso del R/M sistema. Dam metilira na adeninu in igra vlogo pri metiliranju hemimetilirane DNA, ki nastane pri replikaciji.
Na hemimetilirano DNA se na promotorski regiji vežejo DNA vezavni proteini SeqA. Ko je SeqA vezan, preprečuje vezavo Dam na promotorsko regijo in posledično metilacijo samega promotorja. Slednje ima posledico inhibicije ali indukcije transkripcije genov za različne proteine. Spontana disocijacija SeqA dovoli dostop in vezavo Dam na DNA. Ko veže promotor, začne drseti po vijačnici dokler ne pride do specifičnega nukleotidnega zaporedja 5' – GATC – 3' in metilira adenin. Stabilnost adenina v aktivnem mestu omogoči DPPY motiv na proteinu. Po metiliranju promotorja se lahko veže bakterijska dnaA, ki je transkripcijski aktivator [2].
Zgradba metiltransferaz
Naj si bodo nekatere DNA MTaze monomerne, dimerne ali celo oligomerne, so v njih vedno relativno ohranjene domene. Na osnovi funkcije teh domen jih delimo na tri skupine: AdoMet vezavne domene, TRD (target recognition domain) in katalitične domene [2].
Še bolj pa so ohranjeni gradniki teh domen, motivi, ki si sledijo po številčenju od I-X. Čeprav se lahko domene v zaporedju izmenjujejo ali pa se dodajo še druge domene, imajo motivi znotraj domen fiksna zaporedja, kar kaže na vloge teh motivov pri katalitskem procesu.
Katalitična domena objema večinoma motiv IV, ki ima tudi ohranjeno skupno zaporedje. Večinoma se imenuje DPPY, pri čemer se D spreminja glede na različni sev [2,3].
TRD domena prepozna, stabilizira in veže DNA, vendar njeno aminokislinsko zaporedje ni ohranjeno. Pač pa je eno najbolj diverznih zaporedji aminokislinskih ostankov v encimu. Velikokrat so na robu teh zaporedji ponavljajoča se dokaj ohranjena zaporedja. Pomembno je poudariti, da ni direktne povezave med enim aminokislinskim ostankom in enim nukleotidom, marveč so interakcije med TRD in DNA kompleksne narave.
AdoMet vezavna domena se veže na kofaktor. Sestavljena je iz dveh poddomen: ena za vezavo AdoMet in druga za vezavo na prejemnika metilne skupine. AdoMet ima najpomembnejši motiv I. Na osnovi aktivnosti AdoMet vezavne in katalitične domene delimo metiltransferaze R/M sistemov na: eksociklične amino MTaze (N6-adenin MTaza in N4-citozin MTaza) in endociklične MTaze (C5-citozin MTaze) [3].
C5 metilacija
Mehanizem C5 je bil raziskan med prvimi. Posebej dobro preučen je na primeru endonukleaze/metiltransferaze HhaI. Začne se z nukleofilnim napadom C6 atoma v citozinu s tiolno skupino cisteina v Motivu IV. Tvori se kovalentna vez in poveže encim in substrat v kompleks. N3 (na citozinu) se protonira s pomočjo protona, ki ga donira nek kisli aminokislinski ostanek, v primeru HhaI je to glutaminsko kislinski ostanek (E119). To olajša nukleofilni napad. Podre se klasična vezava vodikovih vezi med baznim parom, v katerega je citozin vključen. Hkrati pa se tvorijo vodikove vezi med sedaj glutamatnim ostankom in vodikovim atomom na N4 ter argininom (R165) in kisikom na 2. mestu baznega obroča. Te stabilizirajo položaj baze v katalitičnem mestu. Po potečenem nukleoilnem napadu se sedaj zgodi elektrofilen napad na meso C5, kjer AdoMed svojo metilno skupino, nastane pa AdoHyc. Za zaključek se z napadom baze (najverjetneje vode) odstrani odvečni proton na C5. Prekine pa se tudi C6-S vez preko β eliminacije [1].
Posebnost tega mehanizma je dejstvo, da se zaradi vezave tiolne skupine in C6 atoma celotna baza deformira iz ravnine in zato nima prostora v dvojni vijačnici. To nakazuje na lokacijo katalize reakcije izven dvojne vijačnice [1].
Eksociklične metiltransferaze
Mehanizem, s katerim se prenese metilna skupina na enega od dušikov, vezanih na cikle baz, je dosti manj raziskan, vendar obstaja dokaj verjetna teorija o poteku reakcije.
Nekatere študije so dokazale, da se prenos metilne skupine dogodi neposredno na N6 atomu (M.EcoRI), kar potrjuje, da ni vmesnih intermediatov, kjer bi se prenašala metilna skupina iz enega dušika na drugega. Tako je najbolj splošna razlaga mehanizma, da se dogodi direkten prenos metilne skupine po SN2 reakcijskem mehanizmu z inverzijo simetrije. Podrobnejši vpogled nam pokaže, da se vseeno ustvari intermediat ob prenosu skupine, preden se lahko odcepi proton. Nastane torej N6-metilamonij adeninski kation, ki ga mora katalitsko mesto stabilizirati, da lahko reakcija steče do konca. S pomočjo reševanja M.TaqI struktur v kompleksu z AdoMet je bilo dognano, da v napadu dušika na 6. mestu sodeluje sam dušik najprej kot donor vodikove vezi na aspargin v motivu IV in tudi na kisik skeleta verige med dvema prolinoma v motivu IV. Dušik se delno negativno nabije in ima tako možnost napasti metilno skupino na AdoMet. Nastali N6- metil amonijev intermediat se na koncu deprotonira. To pa privede do končnega produkta N6 -metiliranega adenina [3,4].
Nekaj je tudi predpostavljenih modelov za N4 metiltransferaze. Predvsem za M.PvuII. Domneva je, da se vzpostavijo vodikove vezi med N4 atomom in Ser53 ter Pro54. Hkrati pa je možnost tvorbe van der Waalsovih vezi s Phe56 ostankom (vsi se nahajajo v motivu IV). Aktivacija takega dušika bi lahko potekala po slednjem modelu. Asp96 se poveže z vodikovo vezjo na Ser53. Aktivira se hidroksilna skupina Ser 53 in proton se prenese s citozina preko serina na aspartatno kislinski ostanek. Prenos metilne skupine se dogodi istočasno z deprotonacijo N4. Končno protoniran asp96 tvori vez z N3 na citozinu [3].
Obračanje baz
Pri vseh metiltransferazah je potreba po premestitvi baze, ki se metilira, velika. Razlogi so predvsem sterične narave. Pri C5 metilaciji npr. je to nastanek intermediatov, ki nimajo prostora v klasični B-obliki DNA vijačnice. Tako je bila že prav pri modelu kompleksa HhaI z AdoMet odkrita oblika, kjer je baza, ki je namenjena metilaciji, rotirana iz osi dvojne vijačnice za 180°. Ta baza je tako segala iz malega žleba DNA v katalitično domeno encima. Ista potreba po rotaciji baz se je pokazala pri vseh ostalih vrstah metilaz.
Osrednjo vlogo v sprostitvi baze iz DNA ima TRD. M.TaqI modeli so pokazali, da se ob vezavi struktura vijačnice močno deformira. Razmakne se mali žleb za približno 3 °A, stisne se ogrodje in timidin se pomakne bolj proti centru vijačnice. To je posledica interakcij med skoraj vsemi baznimi pari prepoznavnega zaporedja DNA, razen timidina, ki je par metiliranemu adenozinu. Vse to pripomore prekinitvi van der Waalsovih vezi med tem parom in izpostavitvi adenozina [4].
Metiltransferaze restrikcijsko-modifikacijskih sistemov (R/M sistemi)
Večina bakterijskih DNA metiltransferaz (MTaz) je sklopljenih z restrikcijskim encimom. Skupaj tvorita R/M sisteme, ki ščitijo bakterije pred tujo DNA. MTaza modificira bakterijsko DNA in jo s tem zaščiti pred vezavo restriktaze. Ob vstopu virusne DNA v celico restriktaza nanjo lahko deluje, saj ni modificirana z metilnimi skupinami [1].
R/M sistemi se razdelijo v 4 skupine. Sistem IV dejansko ni pravi RM sistem, saj ne vsebuje MTaze.
Najenostavnejši in tudi najštevilčnejši je sistem II, zanj je značilno, da sta MTaza in restriktaza ločena encima, ki delujeta neodvisno eden od drugega. Prepoznata enako signalno zaporedje na DNA. Večina MTaz sistema II je monomernih proteinov, sestavljenih iz katalitične domene z vezanim AdoMet in TRD domeno. Večinoma prepoznajo kratka palindromska zaporedja na DNA, v dolžini nekje med 2 in 8 nukleotidov. Včasih je prepoznavno zaporedje prekinjeno z nekaj vmesnimi nukleotidi, tako da encim prepozna kratek palindromski odsek, sledi nekaj nesimetričnih nukleotidov in nato spet kratek prepoznavni palindromski odsek. MTaze podsistema IIs prepoznajo nesimetrično zaporedje, zato sta za modifikacijo obeh verig DNA potrebni dve MTazi.
Sistem I je najbolj kompleksen. MTaza in restriktaza sta skupaj s še eno podenoto povezani v heterotrimer. M podenota je odgovorna za metilacijo, R podenota je restriktaza, S podenota pa omogoča vezavo na DNA (vsebuje TRD domeno). Načeloma so vse tri podenote povezane v kompleks, ampak M in S podenota skupaj lahko delujeta neodvisno od R podenote. Vsako podenoto zapisuje svoj gen. Gena za podenoti S in M regulira skupni promotor, medtem ima gen za restriktazo svoj promotor, kar omogoča ločeno regulacijo izražanja. Prepoznavno mesto na DNA je sestavljeno iz dveh krajših asimetričnih zaporedij (3-5 nukleotidov), ki sta ločeni z nekaj vmesnimi nukleotidi. Na vsaki verigi DNA se metilira po en adenin.
R/M sistem III je najmanj raziskana skupina. Podenoti MTaze in restriktaza sestavljata heterodimerni kompleks, za razliko od sistema I pa zraven nimata podenote S. TRD domena se nahaja na obeh ostalih podenotah. Prepoznavno zaporedje je sestavljeno iz 5-6 asimetričnih nukleotidov. Za razliko od preostalih skupin, se metilacija pojavi le na eni verigi DNA, kar pa še vedno omogoča zaščito pred restriktazo.
Hemimetilirana DNA je v vseh primerih zaščitena pred restriktazami, a se v večini primerov metilira tudi na drugi verigi. To je ključnega pomena pri podvojevanju DNA, ko nastaneta 2 hčerinski DNA, ki vsebujeta po eno na novo nastalo verigo in eno verigo iz starševske DNA. Le slednja je metilirana. Če modifikacija na eni sami verigi ne bi bila zadostna zaščita, bi restriktaze prej ali slej razrezale lastne kromosome [1,5].
Regulacija R/M sistemov
1. MTaza se lahko veže na promotor gena za MTazo. S tem prepreči vezavo RNA-polimeraze in prepreči prepisovanje genov.
2. Nekatere MTaze prepoznajo zaporedje, ki je del promotorja za gen MTaze. Metilacija deluje kot represor in gen za MTazo se ne more prepisovati. 1. in 2. način omogočata, da se izražanje MTaze zmanjša, ko je te v celici dovolj. S tem se prepreči, da v primeru pojava virusne DNA restriktaza deluje prej kot MTaza.
3. V primeru prenosa genov za R/M sistem med bakterijami, se mora v bakteriji, ki je prejela gen, najprej izraziti gen za MTazo, šele nato gen za restriktazo. To omogačajo C-proteini (kontrolni proteini). V celici se najprej začne izražati gen za MTazo. Hkrati se začne prepisovati tudi gen za protein C, ki nato aktivira gen za restriktazo. Zamik v nastanku enega in drugega encima je ravno pravšnji, da lahko MTaza modificira kromosome in jih zaščiti. C-protein je homodimerni protein, ki se na palindromsko zaporedje na DNA veže z motivom vijačnica-zavoj-vijačnica [5].
Viri
[1] M. Noyer-Weidner and T. A. Trautner, “Methylation of DNA in Prokaryotes,” in DNA Methylation, Basel: Birkhäuser Basel, 1993, pp. 39–108.
[2] J. Casadesús and D. Low, “Epigenetic gene regulation in the bacterial world.,” Microbiol. Mol. Biol. Rev., vol. 70, no. 3, pp. 830–56, Sep. 2006.
[3] S. Bheemanaik, Y. V. R. Reddy, and D. N. Rao, “Structure, function and mechanism of exocyclic DNA methyltransferases.,” Biochem. J., vol. 399, no. 2, pp. 177–90, Oct. 2006.
[4] W. Gong, M. O’gara, R. M. Blumenthal, X. Cheng, and W. M. Keck, “Structure of PvuII DNA-(cytosine N4) methyltransferase, an example of domain permutation and protein fold assignment,” Oxford University Press, 1997.
[5] G. G. Wilson and N. E. Murray, “Restriction and Modification Systems,” Annu. Rev. Genet., vol. 25, no. 1, pp. 585–627, Dec. 1991.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}