RESHAPE - spreminjanje morfologije nitastih gliv: Difference between revisions
Spelasupej (talk | contribs) No edit summary |
Spelasupej (talk | contribs) No edit summary |
||
| (4 intermediate revisions by the same user not shown) | |||
| Line 14: | Line 14: | ||
=='''Cilji projekta RESHAPE'''== | =='''Cilji projekta RESHAPE'''== | ||
Glavni cilj projekta RESHAPE je bil optimizirati morfologijo nitaste glive ''Aspergillus niger'' s pomočjo pristopov sintezne biologije, kar bi izboljšalo industrijsko proizvodnjo proteinov in drugih malih molekul. Oblikovati so želeli nove signalne peptide, ki bi izboljšali sekrecijo proteinov v ''A. niger''. Poleg tega pa so želeli ustvariti tudi računalniška orodja, ki bi lahko napovedala morfološke vzorce nitastih gliv | Glavni cilj projekta RESHAPE je bil optimizirati morfologijo nitaste glive ''Aspergillus niger'' s pomočjo pristopov sintezne biologije, kar bi izboljšalo industrijsko proizvodnjo proteinov in drugih malih molekul. Oblikovati so želeli nove signalne peptide, ki bi izboljšali sekrecijo proteinov v ''A. niger''. Poleg tega pa so želeli ustvariti tudi računalniška orodja, ki bi lahko na podlagi eksperimentalnih podatkov napovedala morfološke vzorce nitastih gliv. | ||
=='''Spreminjanje morfologije ''Aspergillus niger'' '''== | =='''Spreminjanje morfologije ''Aspergillus niger'' '''== | ||
| Line 31: | Line 31: | ||
=== CRISPR-Cas9 - za spreminjanje genoma === | === CRISPR-Cas9 - za spreminjanje genoma === | ||
Za pripravo novih morfoloških sevov so uporabili tehnologijo CRISPR-Cas9, s katero so izbijali gene iz genoma ''A. niger''. Z USER kloniranjem so oblikovali sedem različnih CRISPR-Cas9 plazmidov, ki si jih je mogoče ogledati na iGEM spletni strani. | Za pripravo novih morfoloških sevov so uporabili tehnologijo CRISPR-Cas9, s katero so izbijali gene iz genoma ''A. niger''. Z USER kloniranjem so oblikovali sedem različnih CRISPR-Cas9 plazmidov, ki si jih je mogoče ogledati na iGEM spletni strani: | ||
-[http://parts.igem.org/wiki/index.php?title=Part:BBa_K3385014 BBa_K3385014], | |||
-[http://parts.igem.org/wiki/index.php?title=Part:BBa_K3385018 BBa_K3385018], | |||
-[http://parts.igem.org/wiki/index.php?title=Part:BBa_K3385022 BBa_K3385022], | |||
-[http://parts.igem.org/wiki/index.php?title=Part:BBa_K3385026 BBa_K3385026], | |||
-[http://parts.igem.org/wiki/index.php?title=Part:BBa_K3385030 BBa_K3385030], | |||
-[http://parts.igem.org/wiki/index.php?title=Part:BBa_K3385034 BBa_K3385034] in | |||
-[http://parts.igem.org/wiki/index.php?title=Part:BBa_K3385038 BBa_K3385038]). | |||
==== Sevi ''A. niger'' z izbitimi geni ==== | ==== Sevi ''A. niger'' z izbitimi geni ==== | ||
| Line 43: | Line 57: | ||
=='''Sekrecija proteinov'''== | =='''Sekrecija proteinov'''== | ||
Nivoje sekrecije so spremljali preko merjenja aktivnosti nativnega encima glukoamilaze (gen ''glaA''). Zapis za protein so najprej izbili iz glivnega genoma, nato pa so ga vanj ponovno integrirali s pomočjo biokock, ki jih je oblikovala danska ekipa leta 2019. Na ta način so zagotovili, da je bila aktivnost encima posledica njihovega inserta in ne naravne produkcije encima. Glukoamilaza je encim, ki razgradi škrob do beta-glukoze. Test sekrecije so naredili tako, da so v supernatant vzorca dodali škrob in po 10 minutni inkubaciji na 37 °C dodali jodov reagent, ki se lahko veže na polimere škorba. Če je prišlo do sekrecije glukoamilaze, je ta škrob razgradila, kar | Nivoje sekrecije so spremljali preko merjenja aktivnosti nativnega encima glukoamilaze (gen ''glaA''). Zapis za protein so najprej izbili iz glivnega genoma, nato pa so ga vanj ponovno integrirali s pomočjo biokock, ki jih je oblikovala danska ekipa leta 2019. Na ta način so zagotovili, da je bila aktivnost encima posledica njihovega inserta in ne naravne produkcije encima. Glukoamilaza je encim, ki razgradi škrob do beta-glukoze. Test sekrecije so naredili tako, da so v supernatant vzorca dodali škrob in po 10 minutni inkubaciji na 37 °C dodali jodov reagent, ki se lahko veže na polimere škorba. Če je prišlo do sekrecije glukoamilaze, je ta škrob razgradila, kar je pomenilo, da je imel jodov reagent na voljo manj škroba za vezavo in je bila zaradi tega sprememba barve manjša. Absorbanco so merili pri 580 nm. | ||
=='''Priprava računalniških modelov'''== | =='''Priprava računalniških modelov'''== | ||
| Line 49: | Line 63: | ||
Za lažjo karakterizacijo novih sevov so razvili tudi dva računalniška modela, ki lahko na podlagi eksperimentalnih podatkov preučujeta morfološke lastnosti ''A. niger''. | Za lažjo karakterizacijo novih sevov so razvili tudi dva računalniška modela, ki lahko na podlagi eksperimentalnih podatkov preučujeta morfološke lastnosti ''A. niger''. | ||
''' | '''Morphologizer''' – je program, ki je sposoben avtomatsko analizirati mikroskopske slike micelijev. Micelije nato tretira kot graf in na podlagi tega določi parametre, kot so razvejanost micelija in kemotropizem. | ||
''' | '''Mycemulator''' – je stohastičen model, ki na podlagi morfoloških parametrov pridobljenih s pomočjo programa Morphologizer in eksperimentalnih podatkov simulira rast micelija. | ||
=='''Rezultati'''== | =='''Rezultati'''== | ||
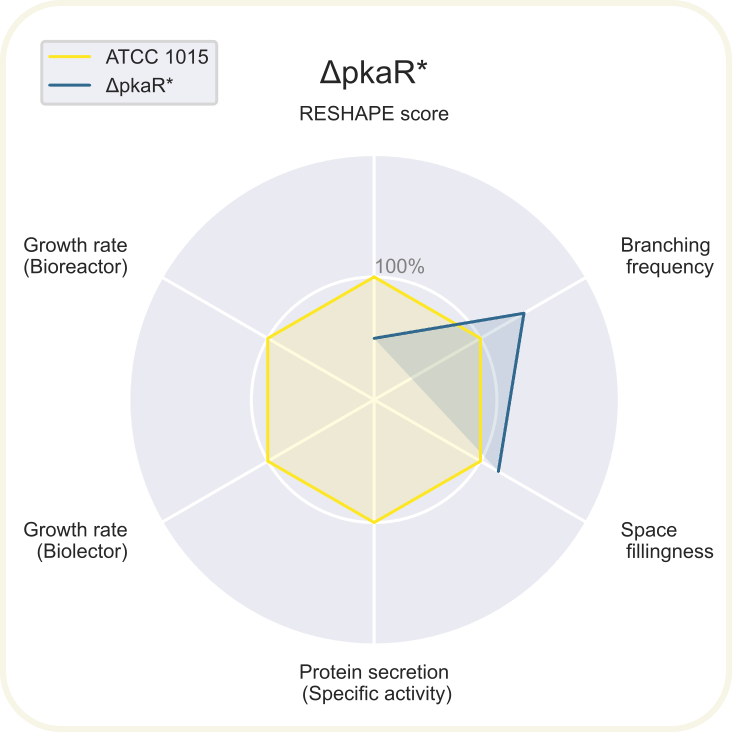
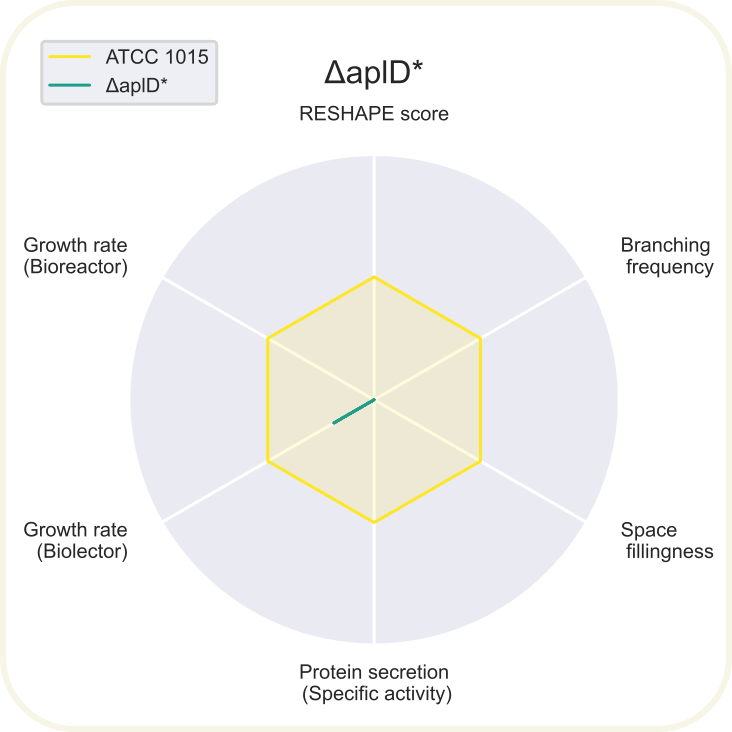
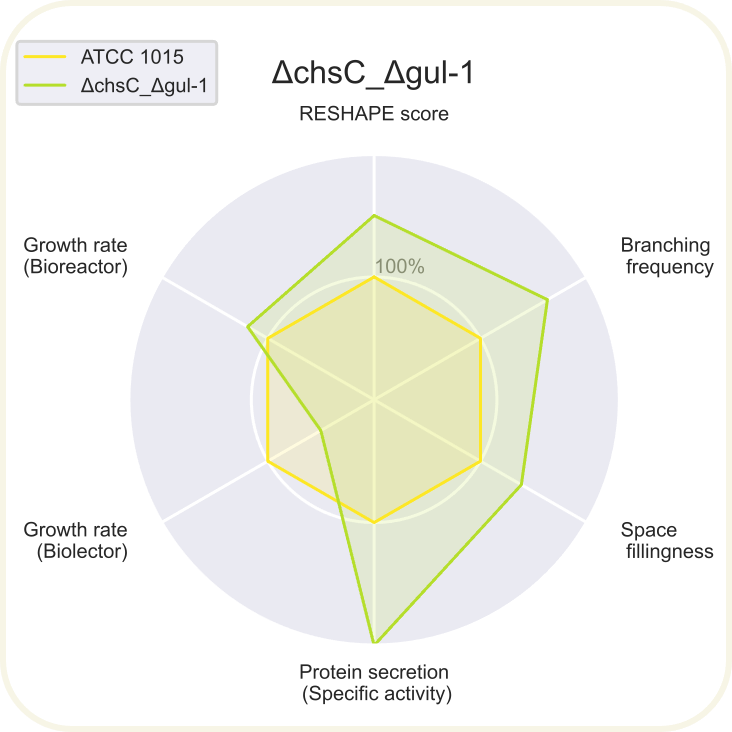
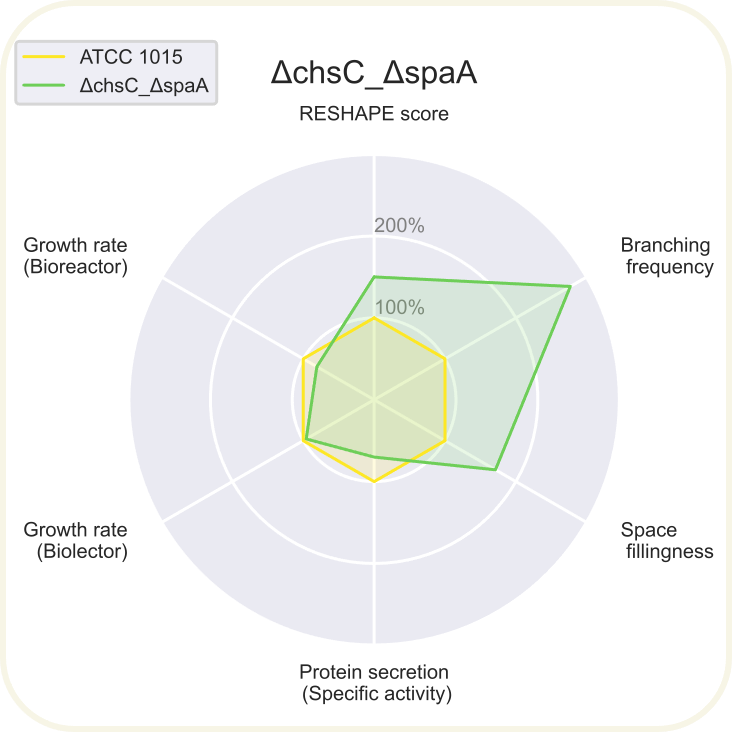
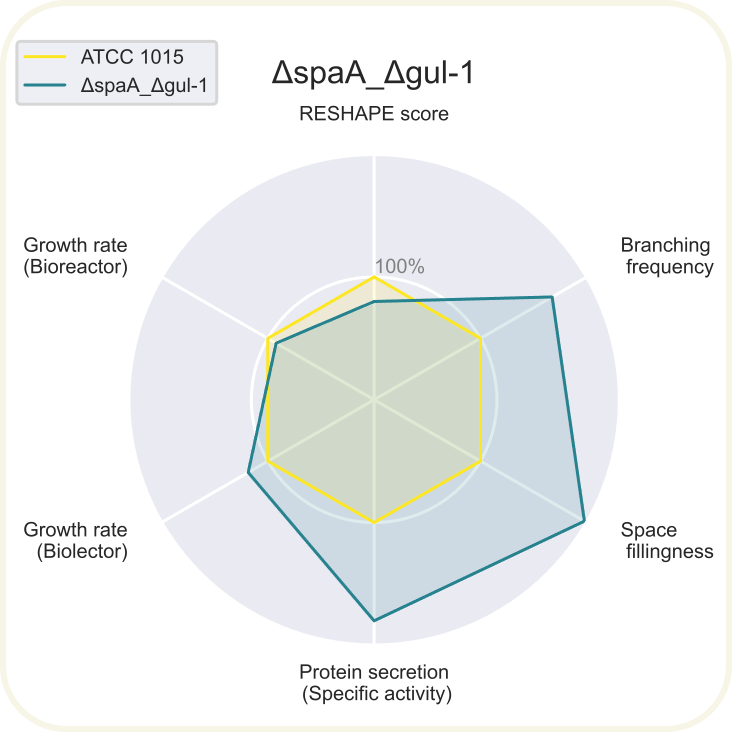
Skupini je uspelo pripraviti devet novih sevov ''A. niger'' z različnimi morfologijami: ''[https://2020.igem.org/wiki/images/d/d3/T--DTU-Denmark--spyder_chsC.png ΔchsC]'', ''[https://2020.igem.org/wiki/images/6/6e/T--DTU-Denmark--spyder_spaA.png ΔspaA]'', ''[https://2020.igem.org/wiki/images/d/de/T--DTU-Denmark--spyder_gul-1.png Δgul-1]'', ''[https://2020.igem.org/wiki/images/c/c8/T--DTU-Denmark--spyder_racA.png ΔracA]'', ''[https://2020.igem.org/wiki/images/0/0a/T--DTU-Denmark--spyder_pkaR.png ΔpkaR]'', ''[https://2020.igem.org/wiki/images/4/44/T--DTU-Denmark--spyder_aplD.png ΔaplD]'', ''[https://2020.igem.org/wiki/images/4/40/T--DTU-Denmark--spyder_chsC_gul-1.png ΔchsC_Δgul-1]'', ''[https://2020.igem.org/wiki/images/6/6d/T--DTU-Denmark--spyder_chsC_spaA.png ΔchsC_ΔspaA]'' in ''[https://2020.igem.org/wiki/images/8/8c/T--DTU-Denmark--spyder_spaA_gul-1.png ΔspaA_Δgul-1]''. Največji potencial je kazal sev z dvema izbitima genoma ''ΔchsC_Δgul-1'', pri | Skupini je uspelo pripraviti devet novih sevov ''A. niger'' z različnimi morfologijami: ''[https://2020.igem.org/wiki/images/d/d3/T--DTU-Denmark--spyder_chsC.png ΔchsC]'', ''[https://2020.igem.org/wiki/images/6/6e/T--DTU-Denmark--spyder_spaA.png ΔspaA]'', ''[https://2020.igem.org/wiki/images/d/de/T--DTU-Denmark--spyder_gul-1.png Δgul-1]'', ''[https://2020.igem.org/wiki/images/c/c8/T--DTU-Denmark--spyder_racA.png ΔracA]'', ''[https://2020.igem.org/wiki/images/0/0a/T--DTU-Denmark--spyder_pkaR.png ΔpkaR]'', ''[https://2020.igem.org/wiki/images/4/44/T--DTU-Denmark--spyder_aplD.png ΔaplD]'', ''[https://2020.igem.org/wiki/images/4/40/T--DTU-Denmark--spyder_chsC_gul-1.png ΔchsC_Δgul-1]'', ''[https://2020.igem.org/wiki/images/6/6d/T--DTU-Denmark--spyder_chsC_spaA.png ΔchsC_ΔspaA]'' in ''[https://2020.igem.org/wiki/images/8/8c/T--DTU-Denmark--spyder_spaA_gul-1.png ΔspaA_Δgul-1]''. Največji potencial je kazal sev z dvema izbitima genoma ''ΔchsC_Δgul-1''. Gen ''chsC'' zapisuje za hitin sintazo, gen ''gul-1'' pa za mRNA vezavni protein, ki sodeluje pri remodeliranju celične stene. Za ta sev so ugotovili, da ima hiperrazvejano morfologijo, kar je bila posledica aditivnega efekta delecije obeh genov. Sev je imel izredno visoko sekrecijo proteinov (kar 200 %) v primerjavi z referenčnim sevom. Sekrecija proteinov je bila najvišja pri sevih z delecijo gena ''[https://2020.igem.org/wiki/images/e/e7/T--DTU-Denmark--all_gluco.png gul-1]''. Oblikovali so tudi plazmide za vstavljanje insertov, s katerimi bi regulirali izražanje, vendar jih zaradi časovnih omejitev niso dokončali in z njimi niso uspeli transformirati gliv. Prav tako jim je uspelo pripraviti knjižnico 11 mutiranih sevov z inserti za različne nativne in sintetične signalne peptide za glukoamilazo, ki so jih določili z orodjem SignalPrepper. Več o pripravi knjižnice si lahko preberete [https://2020.igem.org/Team:DTU-Denmark/Design tukaj]. | ||
=='''Viri in literatura'''== | =='''Viri in literatura'''== | ||
Latest revision as of 10:31, 5 April 2021
RESHAPE je projekt iGEM iz leta 2020, ki so ga naredili študenti iz tehnične univerze DTU na Danskem. Ekipa je želela spremeniti morfologijo nitastih gliv z namenom boljše produkcije proteinov in drugih metabolitov v industriji.
Spletna stran projekta RESHAPE, iGEM 2020 [1]: https://2020.igem.org/Team:DTU-Denmark
Avtorica povzetka: Špela Supej
Uvod
Tako imenovana »bela biotehnologija« je industrijska veja biotehnologije, ki uporablja žive celice kvasovk, gliv, bakterij in rastlin ter njihove encime za sintezo produktov, ki so lahko razgradljivi, njihova produkcija pa zahteva manjšo porabo energije in proizvede manj odpadkov [2]. Nitaste glive predstavljajo veliko večino industrijske produkcije encimov in kemikalij, pri čemer je glavni sev nitastih gliv, v katerem se proizvede okrog 60 % vseh industrijskih encimov, Aspergillus niger [3].
Nitaste glive
Nitaste glive, običajno znane kot plesni, so večcelični organizmi sestavljeni iz struktur, imenovanih hife. Z razraščanjem po organskem materialu oblikujejo večcelična omrežja, ki jih imenujemo miceliji. Glede na genetske in okoljske parametre lahko omrežja zavzamejo različne morfologije, kar omogoča nitastim glivam, da kolonizirajo substrat s čim manjšo količino lastne biomase. Poleg tega imajo izjemne sekretorne sposobnosti, saj lahko preko posebnih organov - hifnih končičev, izločijo kar 30.000 veziklov na minuto [4], [5]. Te lastnosti naredijo nitaste glive izredno zanimive za industrijo, vendar imajo tudi svoje probleme. Hife nitastih gliv povečajo viskoznost v fermentorjih, kar omeji prenos kisika do celic. Te so posledično pod stresom, zaradi česar se zmanjša njihova produktivnost [6]. Izboljšava morfologije nitastih gliv bi imela zaradi tega pozitiven vpliv na industrijsko produkcijo.
Cilji projekta RESHAPE
Glavni cilj projekta RESHAPE je bil optimizirati morfologijo nitaste glive Aspergillus niger s pomočjo pristopov sintezne biologije, kar bi izboljšalo industrijsko proizvodnjo proteinov in drugih malih molekul. Oblikovati so želeli nove signalne peptide, ki bi izboljšali sekrecijo proteinov v A. niger. Poleg tega pa so želeli ustvariti tudi računalniška orodja, ki bi lahko na podlagi eksperimentalnih podatkov napovedala morfološke vzorce nitastih gliv.
Spreminjanje morfologije Aspergillus niger
Morfologija A. niger ima velik vpliv na uspešnost proizvodnje proteinov. Rast v obliki nitastih struktur lahko velikokrat predstavlja težave, saj se filamenti pogosto zavozlajo okrog sestavnih delov fermentacijskih tankov. V skupini iGEM so se zato odločili, da bodo poskušali spremeniti morfologijo A. niger na tak način, da bi bilo delo s sevom olajšano, brez da bi zmanjšali produkcijo proteinov. Identificirali so sedem genov, ki vplivajo na spremembo morfologije in s pomočjo tehnologije CRISPR-Cas9 ustvarili nove seve A. niger s spremenjenimi morfologijami. Geni povezani z morfologijo, ki so jih izbrali, so bili aplD, arfA, chsC, gul-1, pkaR, racA in spaA.
Zakaj so uporabili Aspergillus niger?
Za svoj projekt so izbrali sev nitastih gliv Aspergillus niger, saj se le-ta v industriji najpogosteje uporablja za proizvodnjo encimov in sekundarnih metabolitov. Za delo so uporabili referenčni sev ATCC 1015, ki je imel delecijo ΔkusA, zaradi česar je bilo okvarjeno nehomologno povezovanje koncev. Delecija je sev prisilila v popravljanje prelomov na DNA s pomočjo homologne rekombinacije, kar je omogočilo vstavljanje/deletiranje delov genoma na tarčnih lokacijah. Sev je imel prav tako izbrisan gen pyrG, ki zapisuje za encim orotidin 5'-fosfat dekarboksilazo, zaradi česar ni mogel rasti na gojišču brez prisotnosti uracila oz. uridina, kar so izkoristili kot selekcijski marker [7].
Za izbijanje genov in vstavljanje insercij v genom A. niger so uporabili USER kloniranje in tehniko CRISPR-Cas9.
USER kloniranje - za pripravo plazmidov
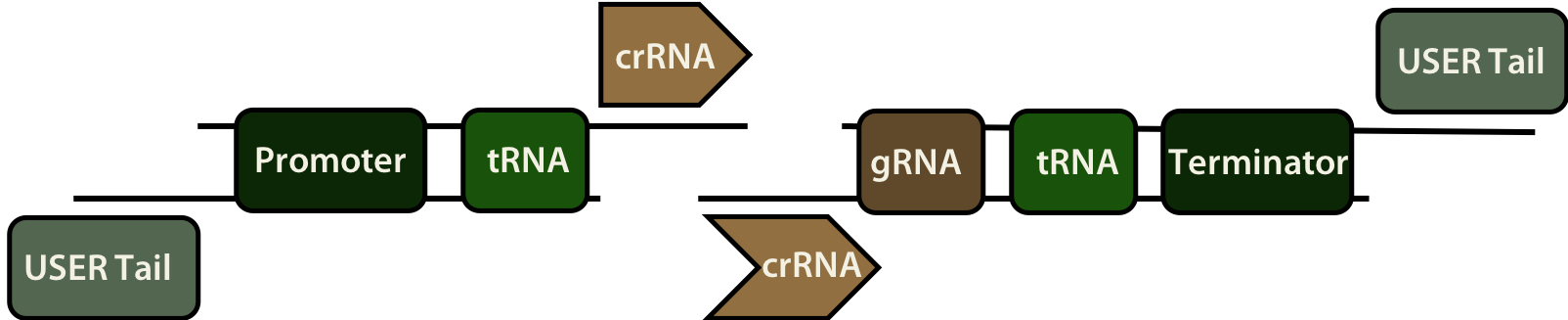
USER kloniranje (ang. Uracil-Specific Excision Reagent) se uporablja za direktno kloniranje insertov v USER kompatibilne vektorje z uporabo USER encima. USER encim združuje encimske aktivnosti uracil DNA glikozilaze – UDG, ki katalizira izrez uracilske baze, in endonukleaze VIII, ki prekine fosfodiestrsko vez na 3' in 5' koncu abazičnega mesta tako, da se deoksiriboza brez baze lahko sprosti iz DNA verige. Encim na ta način ustvari 1 nt dolgo vrzel na mestu uracilskega ostanka [8]. Inserte so ustvarili s pomočjo PCR pomnoževanja začetnih oligonukleotidov, ki so vsebovali en uracil na koncu kompatibilnem z uracilnim klonirnim mestom na vektorju. Produkte so nato obdelali z USER encimom, pri čemer je odstranitev uracila ustvarila previsne konce primerne za kloniranje. USER klonirno kaseto znotraj USER kompatibilnega vektorja so rezali z restrikcijskima encimoma PacI in Nt.BbvCI, ki sta ustvarila previsne konce ujemajoče se s konci pripravljenih insertov. Po mešanju so se tako pripravljeni vektorji in inserti povezali in ustvarili novo oblikovane vektorje, ki so bili primerni za CRISPR-Cas9. Inserti so bili sestavljeni iz promotorja in terminatorja, ki sta zagotovila transkripcijo inserta, usmerjevalne RNA in dveh zaporedij tRNA, ki sta zagotovili samocepitev mRNA po transkripciji in na ta način omogočili vezavo usmerjevalne RNA na protein Cas9.
CRISPR-Cas9 - za spreminjanje genoma
Za pripravo novih morfoloških sevov so uporabili tehnologijo CRISPR-Cas9, s katero so izbijali gene iz genoma A. niger. Z USER kloniranjem so oblikovali sedem različnih CRISPR-Cas9 plazmidov, ki si jih je mogoče ogledati na iGEM spletni strani:
-BBa_K3385034 in
-BBa_K3385038).
Sevi A. niger z izbitimi geni
Če so želeli iz genoma gliv izbrisati specifične gene povezane z morfologijo, so glive transformirali s plazmidom, ki je vseboval tako zapis za Cas9 kot tudi dve sgRNA, kar je omogočilo nastanek dveh zarez (ene navzgor in druge navzdol od tarčnega gena). Glive so bile prav tako transformirane še s popravljalnim oligonukleotidom, ki je bil dolg okoli 90 nt in je deloval kot matrica za homologno rekombinacijo. Kadar so želeli doseči dvojno izbitje genov, so po prvotni transformaciji pridobljene seve ponovno transformirali s CRISPR-Cas9 plazmidom.
Sevi A. niger z zvišano oz. znižano ravnjo izražanja genov
Poleg »knock-out« sevov, so želeli preveriti tudi, kakšen vpliv ima na morfologijo zvišana oz. znižana raven izražanja tarčnih genov. Da bi dobili različne ekspresijske nivoje izražanja genov, so uporabili promotorje skupine DTU iGEM iz leta 2019. Za zvišano raven izražanja so se odločili za močan promotor PLEAPmstA_1 (BBa_K3046005), za znižano pa za šibek promotor PLEAPunk_1 (BBa_K3046006). Za oba promotorja so pripravili biokocke, ki so jih vstavili v plazmide, končni plazmidi pa so nastali, ko so s pomočjo USER kloniranja vanje vstavili še zapise za tarčne gene. Sevom so najprej izbili izbrane gene, nato pa so jih vanje ponovno vstavili z novimi promotorji. Za vstavljanje insercije so glive transformirali z lineariziranim zaporedjem, ki je vsebovalo konstrukt, ki so ga želeli vstaviti v genom, na obeh koncih konstrukta pa so bile 1-2 kb dolge homologne regije. Te regije so bile oblikovane tako, da so omogočale insercijo na specifična mesta znotraj genoma A. niger.
Vpliv morfologije na proizvodnjo proteinov so merili s pomočjo BCA testa (ang. Bicinchoninic Acid Assay), ki omogoča določanje koncentracije proteinov v raztopini. V alkalnem okolju lahko proteini reducirajo Cu2+ do Cu+. Cu+ kationi nato kelirajo dve BCA molekuli, kar vodi do vijoličnega obarvanja. Več proteinov kot je prisotnih, bolj intenzivna bo absorbanca pri 562 nm.
Sekrecija proteinov
Nivoje sekrecije so spremljali preko merjenja aktivnosti nativnega encima glukoamilaze (gen glaA). Zapis za protein so najprej izbili iz glivnega genoma, nato pa so ga vanj ponovno integrirali s pomočjo biokock, ki jih je oblikovala danska ekipa leta 2019. Na ta način so zagotovili, da je bila aktivnost encima posledica njihovega inserta in ne naravne produkcije encima. Glukoamilaza je encim, ki razgradi škrob do beta-glukoze. Test sekrecije so naredili tako, da so v supernatant vzorca dodali škrob in po 10 minutni inkubaciji na 37 °C dodali jodov reagent, ki se lahko veže na polimere škorba. Če je prišlo do sekrecije glukoamilaze, je ta škrob razgradila, kar je pomenilo, da je imel jodov reagent na voljo manj škroba za vezavo in je bila zaradi tega sprememba barve manjša. Absorbanco so merili pri 580 nm.
Priprava računalniških modelov
Za lažjo karakterizacijo novih sevov so razvili tudi dva računalniška modela, ki lahko na podlagi eksperimentalnih podatkov preučujeta morfološke lastnosti A. niger.
Morphologizer – je program, ki je sposoben avtomatsko analizirati mikroskopske slike micelijev. Micelije nato tretira kot graf in na podlagi tega določi parametre, kot so razvejanost micelija in kemotropizem.
Mycemulator – je stohastičen model, ki na podlagi morfoloških parametrov pridobljenih s pomočjo programa Morphologizer in eksperimentalnih podatkov simulira rast micelija.
Rezultati
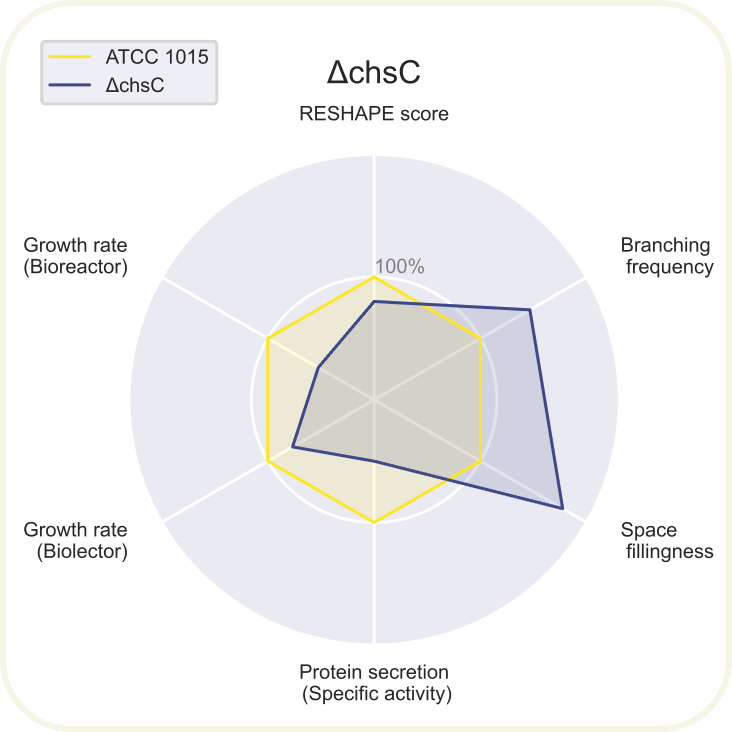
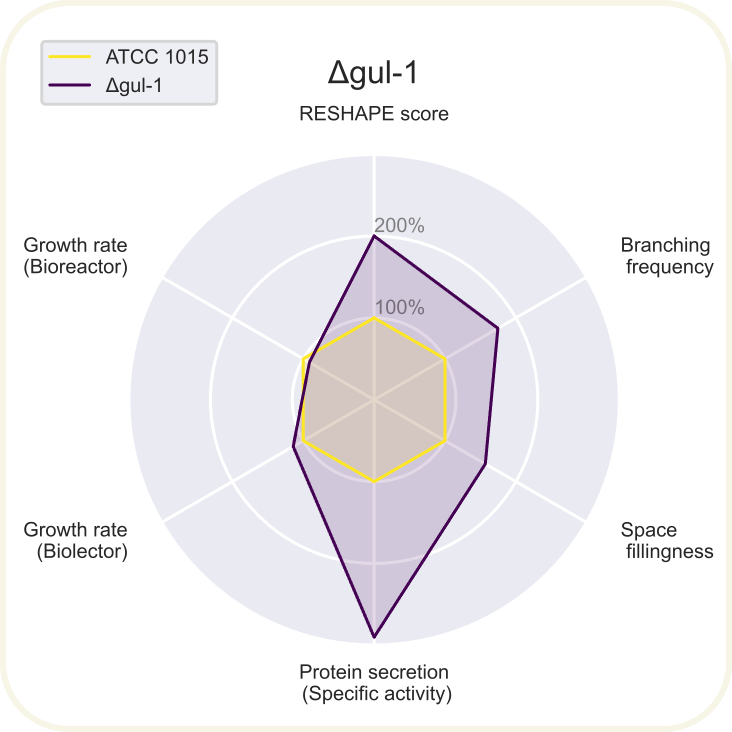
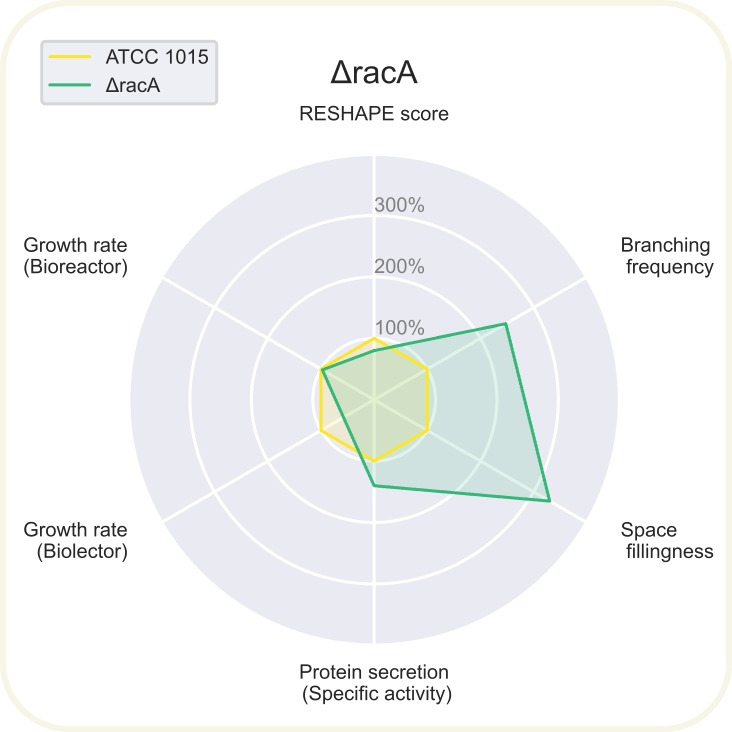
Skupini je uspelo pripraviti devet novih sevov A. niger z različnimi morfologijami: ΔchsC, ΔspaA, Δgul-1, ΔracA, ΔpkaR, ΔaplD, ΔchsC_Δgul-1, ΔchsC_ΔspaA in ΔspaA_Δgul-1. Največji potencial je kazal sev z dvema izbitima genoma ΔchsC_Δgul-1. Gen chsC zapisuje za hitin sintazo, gen gul-1 pa za mRNA vezavni protein, ki sodeluje pri remodeliranju celične stene. Za ta sev so ugotovili, da ima hiperrazvejano morfologijo, kar je bila posledica aditivnega efekta delecije obeh genov. Sev je imel izredno visoko sekrecijo proteinov (kar 200 %) v primerjavi z referenčnim sevom. Sekrecija proteinov je bila najvišja pri sevih z delecijo gena gul-1. Oblikovali so tudi plazmide za vstavljanje insertov, s katerimi bi regulirali izražanje, vendar jih zaradi časovnih omejitev niso dokončali in z njimi niso uspeli transformirati gliv. Prav tako jim je uspelo pripraviti knjižnico 11 mutiranih sevov z inserti za različne nativne in sintetične signalne peptide za glukoamilazo, ki so jih določili z orodjem SignalPrepper. Več o pripravi knjižnice si lahko preberete tukaj.
Viri in literatura
[1] “Team:DTU-Denmark - 2020.igem.org.” https://2020.igem.org/Team:DTU-Denmark (accessed Apr. 02, 2021).
[2] G. Frazzetto, “White biotechnology,” EMBO Rep., vol. 4, no. 9, pp. 835–837, Sep. 2003, doi: 10.1038/sj.embor.embor928.
[3] Siddiqui, S, New and Future Developments in Microbial Biotechnology and Bioengineering. Elsevier, 2016.
[4] V. Meyer et al., “Current challenges of research on filamentous fungi in relation to human welfare and a sustainable bio-economy: a white paper,” Fungal Biol. Biotechnol., vol. 3, no. 1, p. 6, Aug. 2016, doi: 10.1186/s40694-016-0024-8.
[5] G. Steinberg, “Hyphal Growth: a Tale of Motors, Lipids, and the Spitzenkörper,” Eukaryot. Cell, vol. 6, no. 3, pp. 351–360, Mar. 2007, doi: 10.1128/EC.00381-06.
[6] J. Arnau, D. Yaver, and C. M. Hjort, “Strategies and Challenges for the Development of Industrial Enzymes Using Fungal Cell Factories,” in Grand Challenges in Fungal Biotechnology, H. Nevalainen, Ed. Cham: Springer International Publishing, 2020, pp. 179–210.
[7] C. S. Nødvig, J. B. Nielsen, M. E. Kogle, and U. H. Mortensen, “A CRISPR-Cas9 System for Genetic Engineering of Filamentous Fungi,” PLOS ONE, vol. 10, no. 7, p. e0133085, Jul. 2015, doi: 10.1371/journal.pone.0133085.
[8] B. Salomonsen, U. H. Mortensen, and B. A. Halkier, “USER-Derived Cloning Methods and Their Primer Design,” in DNA Cloning and Assembly Methods, vol. 1116, S. Valla and R. Lale, Eds. Totowa, NJ: Humana Press, 2014, pp. 59–72.
[9] G. P. Borin et al., “Comparative Secretome Analysis of Trichoderma reesei and Aspergillus niger during Growth on Sugarcane Biomass,” PLoS ONE, vol. 10, no. 6, Jun. 2015, doi: 10.1371/journal.pone.0129275.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}