Načrtovana pot zvijanja proteinskih origamijev: Difference between revisions
No edit summary |
No edit summary |
||
| (4 intermediate revisions by the same user not shown) | |||
| Line 1: | Line 1: | ||
''Povzeto po članku:'' [https://www.nature.com/articles/s41467-021-21185-5 J. Aupič, Ž. Strmšek, F. Lapenta, D. Pahovnik, T. Pisanski, I. Drobnak, A. Ljubetič, R. Jerala: Designed folding pathway of modular coiled-coil-based proteins. Nat. Commun. 2021, 12(1), str. 1–12.] | ''Povzeto po članku:'' [https://www.nature.com/articles/s41467-021-21185-5 J. Aupič, Ž. Strmšek, F. Lapenta, D. Pahovnik, T. Pisanski, I. Drobnak, A. Ljubetič, R. Jerala: Designed folding pathway of modular coiled-coil-based proteins. Nat. Commun. 2021, 12(1), str. 1–12.] | ||
== ''' | == '''Uvod''' == | ||
Sestavljanje genetskega vezja zgolj iz naravno prisotnih komponent različnih organizmov za vzpostavitev nekaterih sinteznobioloških sistemov ne nujno vedno zadostuje. Želene, vendar v naravi manjkajoče funkcije in biofizikalne lastnosti lahko z modernimi pristopi proteinskega inženirstva do neke mere nadomestimo ali izboljšamo z načrtovanjem dizajnerskih proteinov[1]. Zaradi trenutno aktualnega vse manjšega doprinosa k odkrivanju novih proteinskih zvitij znanstveniki računalniško načrtujejo zvitja in razvijajo nove možnosti, s katerimi bi razširili nabor funkcij makromolekul za uporabo na bionanotehnoloških, sinteznobioloških, strukturnobioloških, biomedicinskih in drugih področjih [2, 3]. Eno večjih ovir pri načrtovanju proteinov ''in silico'' še vedno predstavlja napovedovanje poti zvijanja proteinskih makromolekul [4]. V zadnjem desetletju se veliko pozornosti namenja modularnim nanostrukturam. Razumevanje dinamike zvijanja tovrstnih proteinov nam lahko pomaga pri pojasnjevanju fizikalno-kemijskih mehanizmov, ki usmerjajo zvijanje naravnih modularno zgrajenih proteinov. Ocenjujejo, da modularni proteini predstavljajo petino celotnega evkariontskega proteoma, preučevanje dinamike njihovega zvijanja pa je zaradi velike raznolikosti domen znotraj ene polipeptidne verige pogosto oteženo. Vključeni so v procese celične adhezije in signaliziranja, sodelujejo pa tudi pri sestavljanju znotrajceličnih kompleksov [5]. Napake pri zvijanju modularnih proteinov povezujemo z nekaterimi bolezenskimi stanji [6]. | Sestavljanje genetskega vezja zgolj iz naravno prisotnih komponent različnih organizmov za vzpostavitev nekaterih sinteznobioloških sistemov ne nujno vedno zadostuje. Želene, vendar v naravi manjkajoče funkcije in biofizikalne lastnosti lahko z modernimi pristopi proteinskega inženirstva do neke mere nadomestimo ali izboljšamo z načrtovanjem dizajnerskih proteinov[1]. Zaradi trenutno aktualnega vse manjšega doprinosa k odkrivanju novih proteinskih zvitij znanstveniki računalniško načrtujejo zvitja in razvijajo nove možnosti, s katerimi bi razširili nabor funkcij makromolekul za uporabo na bionanotehnoloških, sinteznobioloških, strukturnobioloških, biomedicinskih in drugih področjih [2, 3]. Eno večjih ovir pri načrtovanju proteinov ''in silico'' še vedno predstavlja napovedovanje poti zvijanja proteinskih makromolekul [4]. V zadnjem desetletju se veliko pozornosti namenja modularnim nanostrukturam. Razumevanje dinamike zvijanja tovrstnih proteinov nam lahko pomaga pri pojasnjevanju fizikalno-kemijskih mehanizmov, ki usmerjajo zvijanje naravnih modularno zgrajenih proteinov. Ocenjujejo, da modularni proteini predstavljajo petino celotnega evkariontskega proteoma, preučevanje dinamike njihovega zvijanja pa je zaradi velike raznolikosti domen znotraj ene polipeptidne verige pogosto oteženo. Vključeni so v procese celične adhezije in signaliziranja, sodelujejo pa tudi pri sestavljanju znotrajceličnih kompleksov [5]. Napake pri zvijanju modularnih proteinov povezujemo z nekaterimi bolezenskimi stanji [6]. | ||
== ''' | == '''Proteinski origamiji''' == | ||
Koncept umetnega sestavljanja osnovnih navidezno samostojnih gradnikov (modulov) v dovršeno tridimenzionalno strukturo se je najprej uveljavil pri oblikovanju nanostruktur nukleinskih kislin. Neposredno parjenje komplementarnih baznih parov (DNA-dupleksi), visoka stabilnost in dostopnost sintetičnih oligonukleotidov poljubnih dolžin so prispevali k relativno enostavnemu in zanesljivemu načrtovanju [https://pubmed.ncbi.nlm.nih.gov/21594298/#&gid=article-figures&pid=fig-4-uid-3 dvo- ] in [https://pubmed.ncbi.nlm.nih.gov/21594298/#&gid=article-figures&pid=fig-6-uid-5 tridimenzionalnih] struktur nukleinskih kislin. Sprva je šlo za samosestavljanje več ločenih verig, kasneje pa se je izkazalo, da je tako s tehnološkega vidika kot tudi ''in vivo'' produkcije struktur nukleinskih kislin ekonomsko smiselneje in enostavneje načrtovati enoverižne konstrukte – t.i. DNA-origamije [7]. | Koncept umetnega sestavljanja osnovnih navidezno samostojnih gradnikov (modulov) v dovršeno tridimenzionalno strukturo se je najprej uveljavil pri oblikovanju nanostruktur nukleinskih kislin. Neposredno parjenje komplementarnih baznih parov (DNA-dupleksi), visoka stabilnost in dostopnost sintetičnih oligonukleotidov poljubnih dolžin so prispevali k relativno enostavnemu in zanesljivemu načrtovanju [https://pubmed.ncbi.nlm.nih.gov/21594298/#&gid=article-figures&pid=fig-4-uid-3 dvo- ] in [https://pubmed.ncbi.nlm.nih.gov/21594298/#&gid=article-figures&pid=fig-6-uid-5 tridimenzionalnih] struktur nukleinskih kislin. Sprva je šlo za samosestavljanje več ločenih verig, kasneje pa se je izkazalo, da je tako s tehnološkega vidika kot tudi ''in vivo'' produkcije struktur nukleinskih kislin ekonomsko smiselneje in enostavneje načrtovati enoverižne konstrukte – t.i. DNA-origamije [7]. | ||
| Line 13: | Line 13: | ||
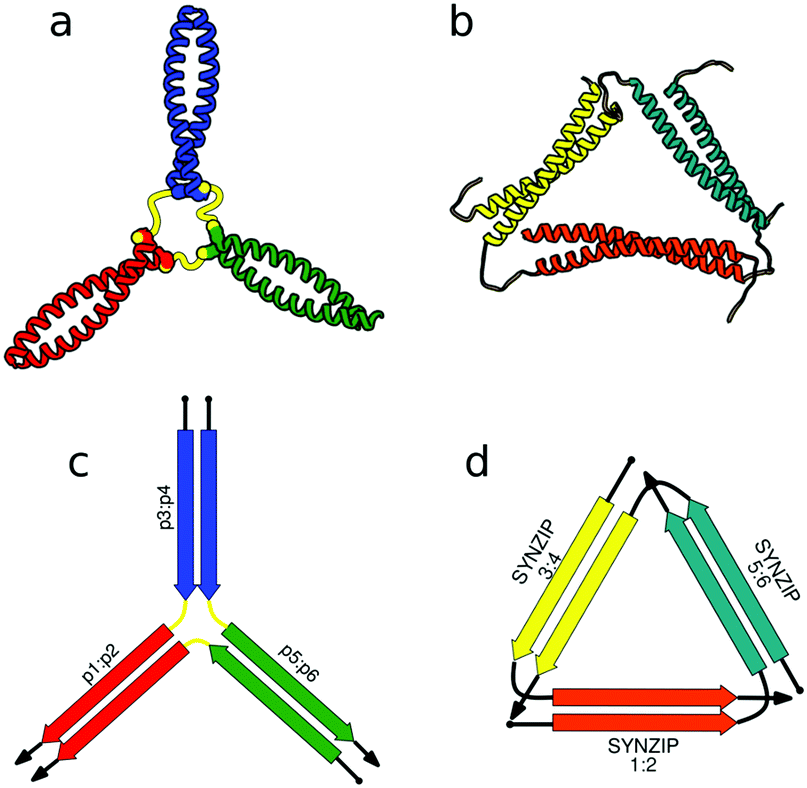
Polipeptidna veriga proteinskega origamija (ang. ''CCPO – coiled-coil protein-origami'') je zgrajena iz medsebojno povezanih dimerov ovitih vijačnic. Vzpostavljanje interakcij med aminokislinskimi ostanki α-vijačnic znotraj posameznega dimera usmerja zvijanje proteina [https://pubs.rsc.org/image/article/2018/CS/c7cs00822h/c7cs00822h-f2_hi-res.gif proti poliedrski strukturi]. Vlogo robov poliedra zavzamejo dimeri ovitih vijačnic [10]. | Polipeptidna veriga proteinskega origamija (ang. ''CCPO – coiled-coil protein-origami'') je zgrajena iz medsebojno povezanih dimerov ovitih vijačnic. Vzpostavljanje interakcij med aminokislinskimi ostanki α-vijačnic znotraj posameznega dimera usmerja zvijanje proteina [https://pubs.rsc.org/image/article/2018/CS/c7cs00822h/c7cs00822h-f2_hi-res.gif proti poliedrski strukturi]. Vlogo robov poliedra zavzamejo dimeri ovitih vijačnic [10]. | ||
=== | ===Motiv ovite vijačnice – osnovni gradnik proteinskega origamija=== | ||
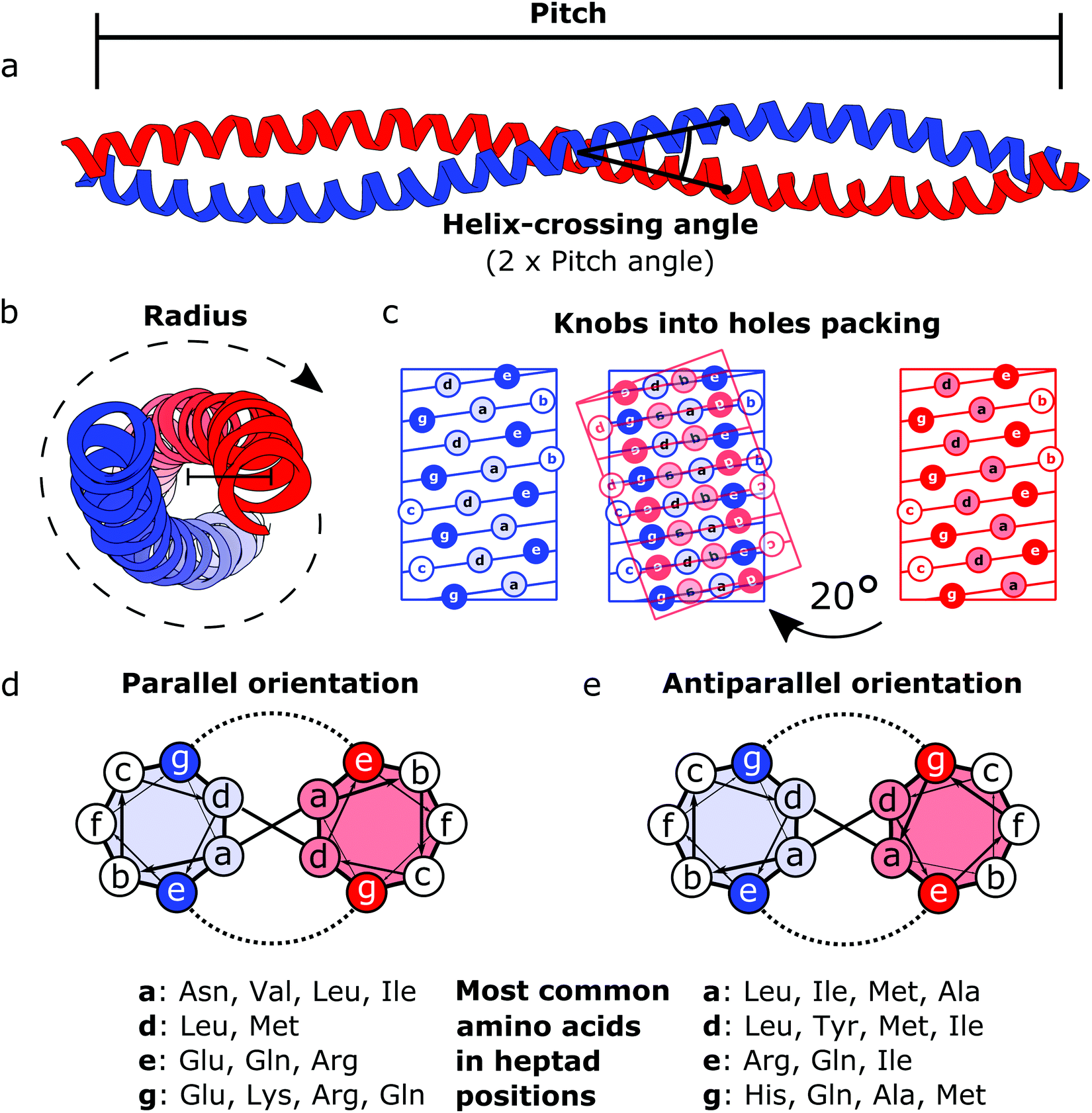
Motiv [https://pubs.rsc.org/image/article/2018/CS/c7cs00822h/c7cs00822h-f1_hi-res.gif ovite vijačnice] je bil za osnovni gradnik proteinskih origamijev izbran predvsem zaradi dobrega poznavanja dinamike njegove poti zvijanja in specifičnosti medsebojnega povezovanja dveh verig. Gre za enega v naravi bolj razširjenih strukturnih proteinskih elementov (predstavlja približno 10 % evkariontskega proteoma), katerih osrednja funkcija je posredovanje interakcij protein-protein in protein-nukleinska kislina. Podobno funkcijo lahko poleg strukturne pričakujemo tudi od načrtovanih proteinskih origamijev. Za motiv je značilno paralelno ali antiparalelno medsebojno ovijanje dveh α-vijačnic po principu »gumbkov v luknjice« (ang. ''knobs into holes''). Da pride do vzpostavitve ustreznih hidrofobnih in polarnih stabilizirajočih interakcij, mora biti v aminokislinskem zaporedju izpolnjen pogoj heptadnih ponovitev [10]. | Motiv [https://pubs.rsc.org/image/article/2018/CS/c7cs00822h/c7cs00822h-f1_hi-res.gif ovite vijačnice] je bil za osnovni gradnik proteinskih origamijev izbran predvsem zaradi dobrega poznavanja dinamike njegove poti zvijanja in specifičnosti medsebojnega povezovanja dveh verig. Gre za enega v naravi bolj razširjenih strukturnih proteinskih elementov (predstavlja približno 10 % evkariontskega proteoma), katerih osrednja funkcija je posredovanje interakcij protein-protein in protein-nukleinska kislina. Podobno funkcijo lahko poleg strukturne pričakujemo tudi od načrtovanih proteinskih origamijev. Za motiv je značilno paralelno ali antiparalelno medsebojno ovijanje dveh α-vijačnic po principu »gumbkov v luknjice« (ang. ''knobs into holes''). Da pride do vzpostavitve ustreznih hidrofobnih in polarnih stabilizirajočih interakcij, mora biti v aminokislinskem zaporedju izpolnjen pogoj heptadnih ponovitev [10]. | ||
=== | ===Nizi ovitih vijačnic, ki so medsebojno pravokotne=== | ||
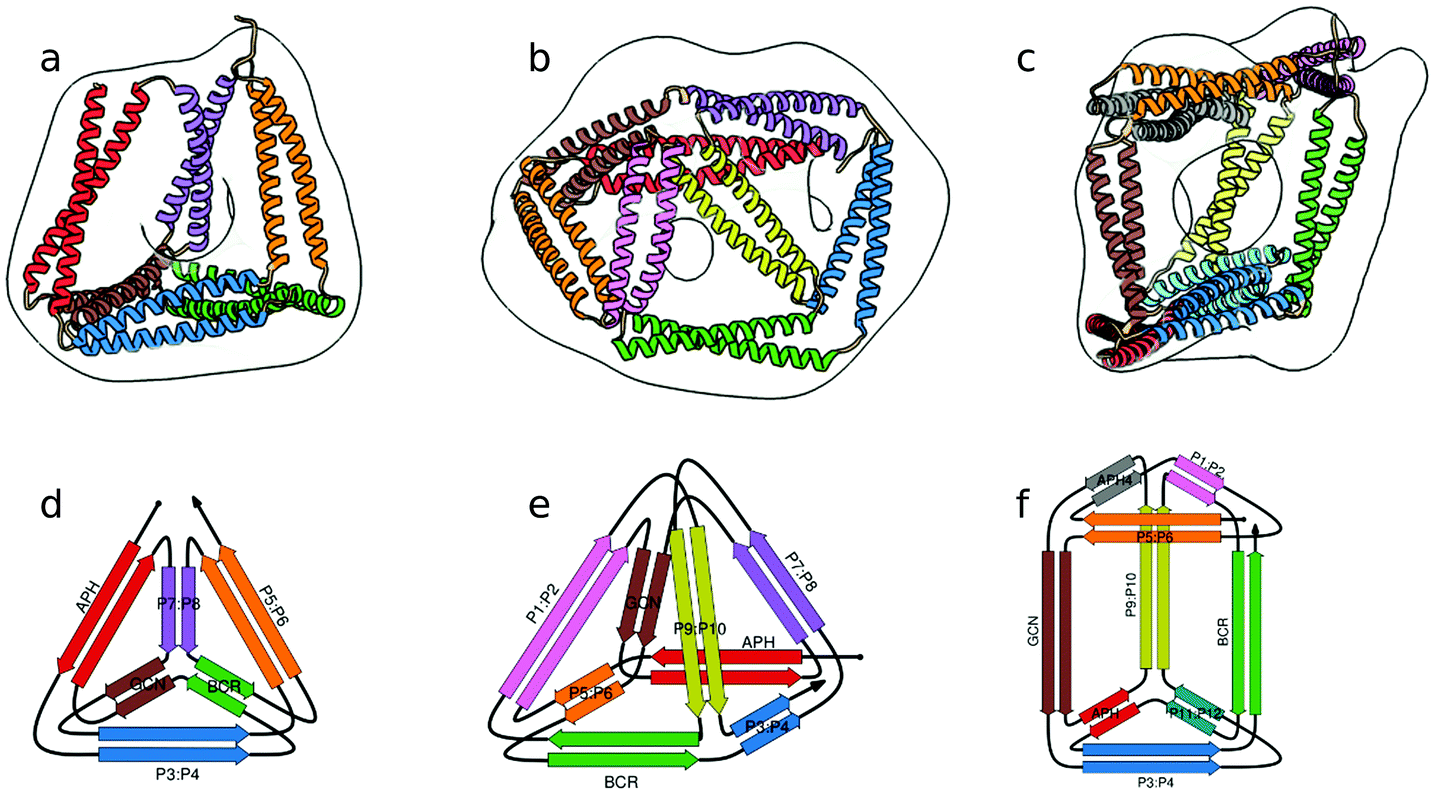
Proteinske kletke so načrtovali na podlagi niza pravokotnih ovitih vijačnic. Zanj je značilno, da posamezne α-vijačnice interagirajo izključno z eno vnaprej predvideno partnersko α-vijačnico. Do interakcij z drugimi α-vijačnicami (ang. ''cross-interact'') v nizu ne pride. Med posamezne osnovne gradnike so v zaporedje strateško vstavili glicinske vmesnike, kar je tudi eden glavnih razlogov, da pri zvijanju pride do želene ortogonalne prostorske razporeditve osnovnih gradnikov. Razvitih je bilo več nizov, ki so tvorili različno velike proteinske kletke – [https://pubs.rsc.org/image/article/2018/CS/c7cs00822h/c7cs00822h-f3_hi-res.gif tetraeder, kvadratna piramida, trigonalna prizma] [10] … | Proteinske kletke so načrtovali na podlagi niza pravokotnih ovitih vijačnic. Zanj je značilno, da posamezne α-vijačnice interagirajo izključno z eno vnaprej predvideno partnersko α-vijačnico. Do interakcij z drugimi α-vijačnicami (ang. ''cross-interact'') v nizu ne pride. Med posamezne osnovne gradnike so v zaporedje strateško vstavili glicinske vmesnike, kar je tudi eden glavnih razlogov, da pri zvijanju pride do želene ortogonalne prostorske razporeditve osnovnih gradnikov. Razvitih je bilo več nizov, ki so tvorili različno velike proteinske kletke – [https://pubs.rsc.org/image/article/2018/CS/c7cs00822h/c7cs00822h-f3_hi-res.gif tetraeder, kvadratna piramida, trigonalna prizma] [10] … | ||
==''' | =='''Sledenje zvijanju proteinskih origamijev'''== | ||
Tekom zvijanja lahko pri naravnih proteinih s tandemskimi ponovitvami določenih elementov pride do t.i. izmenjave domen znotraj polipeptidne verige (ang. ''domain-swapping''), česar posledica je stabilizacija nenativne strukture, ki je lahko zaradi tvorjenja netopnih agregatov patološka [4]. | Tekom zvijanja lahko pri naravnih proteinih s tandemskimi ponovitvami določenih elementov pride do t.i. izmenjave domen znotraj polipeptidne verige (ang. ''domain-swapping''), česar posledica je stabilizacija nenativne strukture, ki je lahko zaradi tvorjenja netopnih agregatov patološka [4]. | ||
| Line 25: | Line 25: | ||
S simulacijami molekulske dinamike so za številne predhodno sintetizirane tetraedrične proteinske origamije določili optimalno lego ponavljajočih se dimernih vijačnih modulov. Analiza trajektorij molekulske dinamike je pokazala, da procesa zvijanja posameznih modulov in dimerizacija dveh α-vijačnic potekata neodvisno od drugih (v primeru, ko proteinski origami ne vsebuje identičnih modulov). Da je bil izpolnjen pogoj formiranosti motiva ovite vijačnice, je moralo biti vzpostavljenih vsaj 50 % interakcij, prisotnih v nativnem stanju. Simulacije molekulske dinamike so pokazale, da bodo verige, ki se v prostoru nahajajo bližje skupaj, dimerizirale z večjo verjetnostjo. To so eksperimentalno potrdile tudi [https://www.nature.com/articles/s41467-021-21185-5/figures/2 biofizikalne meritve] stabilnosti proteinskih origamijev [4]. | S simulacijami molekulske dinamike so za številne predhodno sintetizirane tetraedrične proteinske origamije določili optimalno lego ponavljajočih se dimernih vijačnih modulov. Analiza trajektorij molekulske dinamike je pokazala, da procesa zvijanja posameznih modulov in dimerizacija dveh α-vijačnic potekata neodvisno od drugih (v primeru, ko proteinski origami ne vsebuje identičnih modulov). Da je bil izpolnjen pogoj formiranosti motiva ovite vijačnice, je moralo biti vzpostavljenih vsaj 50 % interakcij, prisotnih v nativnem stanju. Simulacije molekulske dinamike so pokazale, da bodo verige, ki se v prostoru nahajajo bližje skupaj, dimerizirale z večjo verjetnostjo. To so eksperimentalno potrdile tudi [https://www.nature.com/articles/s41467-021-21185-5/figures/2 biofizikalne meritve] stabilnosti proteinskih origamijev [4]. | ||
==''' | ==='''Matematični model za načrtovanje zvijanja proteinskih origamijev'''=== | ||
Na podlagi ugotovljenega so razvili matematični model, ki upošteva hitrost zvijanja glede na razdaljo med posameznimi elementi. Z njim bi bilo z umeščanjem strukturnega elementa na natančno predvideno mesto v aminokislinsko zaporedje mogoče oblikovati proteinske origamije, katerih pot zvijanja bi lahko vnaprej zanesljivo predvideli. Po modelu se posamezna vijačnica v vsakem trenutku zvijanja pari s sebi najbližjo kompatibilno vijačnico. Model torej upošteva splošno uveljavljen princip, ki ga pojasnjujejo številne eksperimentalne in teoretične raziskave – da je zvijanje proteina v prvi vrsti odvisno od topologije njegovih elementov. V primeru, da je v nekem trenutku razdalja ene vijačnice do dveh drugih enaka, model to predvidi in v nadaljevanju pri izračunu upošteva vse možne poti zvijanja [4]. | Na podlagi ugotovljenega so razvili matematični model, ki upošteva hitrost zvijanja glede na razdaljo med posameznimi elementi. Z njim bi bilo z umeščanjem strukturnega elementa na natančno predvideno mesto v aminokislinsko zaporedje mogoče oblikovati proteinske origamije, katerih pot zvijanja bi lahko vnaprej zanesljivo predvideli. Po modelu se posamezna vijačnica v vsakem trenutku zvijanja pari s sebi najbližjo kompatibilno vijačnico. Model torej upošteva splošno uveljavljen princip, ki ga pojasnjujejo številne eksperimentalne in teoretične raziskave – da je zvijanje proteina v prvi vrsti odvisno od topologije njegovih elementov. V primeru, da je v nekem trenutku razdalja ene vijačnice do dveh drugih enaka, model to predvidi in v nadaljevanju pri izračunu upošteva vse možne poti zvijanja [4]. | ||
Za opis verjetnosti nekega proteinskega origamija, da se bo pravilno zvil, so uvedli koeficient P<sub>F</sub>, ki predstavlja razmerje med številom poti zvijanja, ki vodijo do poliedrične končne strukture, in številom vseh možnih poti zvijanja. Za polipeptidne verige, ki so sestavljene iz različnih modulov je vrednost koeficienta P<sub>F</sub> enaka 1, medtem ko pomnožitev istega modula znotraj iste polipeptidne verige proteinskega origamija ta koeficient zmanjša. Efektivno se to kaže v večji verjetnosti neustrezno zvitih stanj (stanj, ki ne izkazujejo lastnosti poliedra). Da bi ugotovili, koliko identičnih modulov lahko uvedejo, da še ne pride do napačnega zvijanja, so pripravili tetraedrične proteinske origamije z dvema, uspela pa jim je tudi priprava s tremi identičnimi moduli od celokupno šestih, ki sestavljajo tetraedrično proteinsko kletko. Za uspešnost zvijanja je bilo ključnega pomena, ali so bili identični moduli sosednji. [https://www.nature.com/articles/s41467-021-21185-5/figures/4 Rezultati] so potrdili hipotezo, da kopičenje različnih motivov v polipeptidno verigo zmanjšuje verjetnost agregacije proteinov [4]. | Za opis verjetnosti nekega proteinskega origamija, da se bo pravilno zvil, so uvedli koeficient P<sub>F</sub>, ki predstavlja razmerje med številom poti zvijanja, ki vodijo do poliedrične končne strukture, in številom vseh možnih poti zvijanja. Za polipeptidne verige, ki so sestavljene iz različnih modulov, je vrednost koeficienta P<sub>F</sub> enaka 1, medtem ko pomnožitev istega modula znotraj iste polipeptidne verige proteinskega origamija ta koeficient zmanjša. Efektivno se to kaže v večji verjetnosti neustrezno zvitih stanj (stanj, ki ne izkazujejo lastnosti poliedra). Da bi ugotovili, koliko identičnih modulov lahko uvedejo, da še ne pride do napačnega zvijanja, so pripravili tetraedrične proteinske origamije z dvema, uspela pa jim je tudi priprava s tremi identičnimi moduli od celokupno šestih, ki sestavljajo tetraedrično proteinsko kletko. Za uspešnost zvijanja je bilo ključnega pomena, ali so bili identični moduli sosednji. [https://www.nature.com/articles/s41467-021-21185-5/figures/4 Rezultati] so potrdili hipotezo, da kopičenje različnih motivov v polipeptidno verigo zmanjšuje verjetnost agregacije proteinov [4]. | ||
==''' | =='''Zaključek'''== | ||
Raziskava med drugim prispeva k potrditvi, da zvijanje proteinskih molekul ne poteče usklajeno po korakih zgolj ene poti, hkrati pa tudi z vzpostavitvijo modela za napovedovanje zvijanja nakazuje, da teh poti ni neobvladljivo veliko. Slednja srednja pot mehanizma zvijanja proteinov, ki predvideva favoriziranje omejenega števila možnih vzporednih stopenj zvijanja, je bila predhodno nakazana že z drugimi študijami. Zaradi poudarjene modularnosti, ki je posledica specifične topološke zgradbe proteinskih origamijev, ti predstavljajo zanimiv in radikalno preprost model za nadaljnje študije zvijanja večjih modularnih proteinov. Za oceno razdalje med interagirajočima elementoma ovitih vijačnic so uporabili molekulsko dinamiko in eksperimentalne meritve kinetike zvijanja. Na podlagi rezultatov so razvili matematični model za napovedovanje verjetnosti zvijanja proteinskih origamijev z različnim številom osnovnih gradnikov. | Raziskava med drugim prispeva k potrditvi, da zvijanje proteinskih molekul ne poteče usklajeno po korakih zgolj ene poti, hkrati pa tudi z vzpostavitvijo modela za napovedovanje zvijanja nakazuje, da teh poti ni neobvladljivo veliko. Slednja srednja pot mehanizma zvijanja proteinov, ki predvideva favoriziranje omejenega števila možnih vzporednih stopenj zvijanja, je bila predhodno nakazana že z drugimi študijami. Zaradi poudarjene modularnosti, ki je posledica specifične topološke zgradbe proteinskih origamijev, ti predstavljajo zanimiv in radikalno preprost model za nadaljnje študije zvijanja večjih modularnih proteinov. Za oceno razdalje med interagirajočima elementoma ovitih vijačnic so uporabili molekulsko dinamiko in eksperimentalne meritve kinetike zvijanja. Na podlagi rezultatov so razvili matematični model za napovedovanje verjetnosti zvijanja proteinskih origamijev z različnim številom osnovnih gradnikov. | ||
Proteinski origamiji pa niso zanimivi zgolj z vidika preučevanja njihove dinamike zvijanja. Zaradi svoje značilne strukture in lastnosti (prilagodljivost, biorazgradljivost, sposobnost samosestavljanja, enostavnost za manipulacijo) imajo velik potencial v smeri razvoja biomedicinskih materialov, npr. kot dostavni sistemi za terapevtske učinkovine ali kot ogrodje za nekatera cepiva. Gre namreč za | Proteinski origamiji pa niso zanimivi zgolj z vidika preučevanja njihove dinamike zvijanja. Zaradi svoje značilne strukture in lastnosti (prilagodljivost, biorazgradljivost, sposobnost samosestavljanja, enostavnost za manipulacijo) imajo velik potencial v smeri razvoja biomedicinskih materialov, npr. kot dostavni sistemi za terapevtske učinkovine ali kot ogrodje za nekatera cepiva. Gre namreč za edini do sedaj poznani tip strukture proteinske kletke s središčno praznino, ki je sestavljen zgolj iz ene polipeptidne verige [10]. | ||
== ''' | == '''Viri'''== | ||
[1.] P. Gainza-Cirauqui, B. E. Correia: Computational protein design — the next generation tool to expand synthetic biology applications. Curr. ''Opin. Biotechnol.'' '''2018''', 52, str. 145–152. | [1.] P. Gainza-Cirauqui, B. E. Correia: Computational protein design — the next generation tool to expand synthetic biology applications. Curr. ''Opin. Biotechnol.'' '''2018''', 52, str. 145–152. | ||
Latest revision as of 06:39, 13 April 2021
Uvod
Sestavljanje genetskega vezja zgolj iz naravno prisotnih komponent različnih organizmov za vzpostavitev nekaterih sinteznobioloških sistemov ne nujno vedno zadostuje. Želene, vendar v naravi manjkajoče funkcije in biofizikalne lastnosti lahko z modernimi pristopi proteinskega inženirstva do neke mere nadomestimo ali izboljšamo z načrtovanjem dizajnerskih proteinov[1]. Zaradi trenutno aktualnega vse manjšega doprinosa k odkrivanju novih proteinskih zvitij znanstveniki računalniško načrtujejo zvitja in razvijajo nove možnosti, s katerimi bi razširili nabor funkcij makromolekul za uporabo na bionanotehnoloških, sinteznobioloških, strukturnobioloških, biomedicinskih in drugih področjih [2, 3]. Eno večjih ovir pri načrtovanju proteinov in silico še vedno predstavlja napovedovanje poti zvijanja proteinskih makromolekul [4]. V zadnjem desetletju se veliko pozornosti namenja modularnim nanostrukturam. Razumevanje dinamike zvijanja tovrstnih proteinov nam lahko pomaga pri pojasnjevanju fizikalno-kemijskih mehanizmov, ki usmerjajo zvijanje naravnih modularno zgrajenih proteinov. Ocenjujejo, da modularni proteini predstavljajo petino celotnega evkariontskega proteoma, preučevanje dinamike njihovega zvijanja pa je zaradi velike raznolikosti domen znotraj ene polipeptidne verige pogosto oteženo. Vključeni so v procese celične adhezije in signaliziranja, sodelujejo pa tudi pri sestavljanju znotrajceličnih kompleksov [5]. Napake pri zvijanju modularnih proteinov povezujemo z nekaterimi bolezenskimi stanji [6].
Proteinski origamiji
Koncept umetnega sestavljanja osnovnih navidezno samostojnih gradnikov (modulov) v dovršeno tridimenzionalno strukturo se je najprej uveljavil pri oblikovanju nanostruktur nukleinskih kislin. Neposredno parjenje komplementarnih baznih parov (DNA-dupleksi), visoka stabilnost in dostopnost sintetičnih oligonukleotidov poljubnih dolžin so prispevali k relativno enostavnemu in zanesljivemu načrtovanju dvo- in tridimenzionalnih struktur nukleinskih kislin. Sprva je šlo za samosestavljanje več ločenih verig, kasneje pa se je izkazalo, da je tako s tehnološkega vidika kot tudi in vivo produkcije struktur nukleinskih kislin ekonomsko smiselneje in enostavneje načrtovati enoverižne konstrukte – t.i. DNA-origamije [7].
Znanje robustnih strategij načrtovanja DNA-origamijev je v kombinaciji z raznolikostjo, ki jo ponuja aminokislinska sestava polipeptidne verige, sčasoma privedlo do snovanja poliedrskih proteinskih kletk. Vlogo DNA-dupleksa – značilnega osnovnega gradnika DNA-origamijev – je R. Jerala s sod. pri načrtovanju proteinskih kletk zamenjal z dimernim proteinskim motivom ovite vijačnice. Proteinske kletke, sestavljene iz medsebojno pravokotnih ovitih vijačnic kot osnovnih gradnikov, je raziskovalna skupina poimenovala proteinski origamiji [8].
Origami« izvira iz poimenovanja za japonsko umetnost zvijanja papirja in ponazarja analogijo z zvijanjem načrtovanih modularno sestavljenih makromolekul – proteinov in nukleinskih kislin [9].
Polipeptidna veriga proteinskega origamija (ang. CCPO – coiled-coil protein-origami) je zgrajena iz medsebojno povezanih dimerov ovitih vijačnic. Vzpostavljanje interakcij med aminokislinskimi ostanki α-vijačnic znotraj posameznega dimera usmerja zvijanje proteina proti poliedrski strukturi. Vlogo robov poliedra zavzamejo dimeri ovitih vijačnic [10].
Motiv ovite vijačnice – osnovni gradnik proteinskega origamija
Motiv ovite vijačnice je bil za osnovni gradnik proteinskih origamijev izbran predvsem zaradi dobrega poznavanja dinamike njegove poti zvijanja in specifičnosti medsebojnega povezovanja dveh verig. Gre za enega v naravi bolj razširjenih strukturnih proteinskih elementov (predstavlja približno 10 % evkariontskega proteoma), katerih osrednja funkcija je posredovanje interakcij protein-protein in protein-nukleinska kislina. Podobno funkcijo lahko poleg strukturne pričakujemo tudi od načrtovanih proteinskih origamijev. Za motiv je značilno paralelno ali antiparalelno medsebojno ovijanje dveh α-vijačnic po principu »gumbkov v luknjice« (ang. knobs into holes). Da pride do vzpostavitve ustreznih hidrofobnih in polarnih stabilizirajočih interakcij, mora biti v aminokislinskem zaporedju izpolnjen pogoj heptadnih ponovitev [10].
Nizi ovitih vijačnic, ki so medsebojno pravokotne
Proteinske kletke so načrtovali na podlagi niza pravokotnih ovitih vijačnic. Zanj je značilno, da posamezne α-vijačnice interagirajo izključno z eno vnaprej predvideno partnersko α-vijačnico. Do interakcij z drugimi α-vijačnicami (ang. cross-interact) v nizu ne pride. Med posamezne osnovne gradnike so v zaporedje strateško vstavili glicinske vmesnike, kar je tudi eden glavnih razlogov, da pri zvijanju pride do želene ortogonalne prostorske razporeditve osnovnih gradnikov. Razvitih je bilo več nizov, ki so tvorili različno velike proteinske kletke – tetraeder, kvadratna piramida, trigonalna prizma [10] …
Sledenje zvijanju proteinskih origamijev
Tekom zvijanja lahko pri naravnih proteinih s tandemskimi ponovitvami določenih elementov pride do t.i. izmenjave domen znotraj polipeptidne verige (ang. domain-swapping), česar posledica je stabilizacija nenativne strukture, ki je lahko zaradi tvorjenja netopnih agregatov patološka [4].
Cilj raziskave je bilo ugotoviti, ali lahko pri načrtovanju proteinskih origamijev uporabijo več enakih dimerov ovitih vijačnic, ne da bi pri tem prišlo do izmenjave domen, ter tako poenostavijo njihovo načrtovanje. Uporaba identičnih modulov pri načrtovanju proteinskih origamijev bi bila mogoča v primeru, da bi bilo proteinu v procesu zvijanja omogočeno ustrezno parjenje domen, kar pa bi bilo mogoče, če bi lahko pot zvijanja proteinskih origamijev in sestavljanja posameznih domen natančno načrtovali [4].
S simulacijami molekulske dinamike so za številne predhodno sintetizirane tetraedrične proteinske origamije določili optimalno lego ponavljajočih se dimernih vijačnih modulov. Analiza trajektorij molekulske dinamike je pokazala, da procesa zvijanja posameznih modulov in dimerizacija dveh α-vijačnic potekata neodvisno od drugih (v primeru, ko proteinski origami ne vsebuje identičnih modulov). Da je bil izpolnjen pogoj formiranosti motiva ovite vijačnice, je moralo biti vzpostavljenih vsaj 50 % interakcij, prisotnih v nativnem stanju. Simulacije molekulske dinamike so pokazale, da bodo verige, ki se v prostoru nahajajo bližje skupaj, dimerizirale z večjo verjetnostjo. To so eksperimentalno potrdile tudi biofizikalne meritve stabilnosti proteinskih origamijev [4].
Matematični model za načrtovanje zvijanja proteinskih origamijev
Na podlagi ugotovljenega so razvili matematični model, ki upošteva hitrost zvijanja glede na razdaljo med posameznimi elementi. Z njim bi bilo z umeščanjem strukturnega elementa na natančno predvideno mesto v aminokislinsko zaporedje mogoče oblikovati proteinske origamije, katerih pot zvijanja bi lahko vnaprej zanesljivo predvideli. Po modelu se posamezna vijačnica v vsakem trenutku zvijanja pari s sebi najbližjo kompatibilno vijačnico. Model torej upošteva splošno uveljavljen princip, ki ga pojasnjujejo številne eksperimentalne in teoretične raziskave – da je zvijanje proteina v prvi vrsti odvisno od topologije njegovih elementov. V primeru, da je v nekem trenutku razdalja ene vijačnice do dveh drugih enaka, model to predvidi in v nadaljevanju pri izračunu upošteva vse možne poti zvijanja [4].
Za opis verjetnosti nekega proteinskega origamija, da se bo pravilno zvil, so uvedli koeficient PF, ki predstavlja razmerje med številom poti zvijanja, ki vodijo do poliedrične končne strukture, in številom vseh možnih poti zvijanja. Za polipeptidne verige, ki so sestavljene iz različnih modulov, je vrednost koeficienta PF enaka 1, medtem ko pomnožitev istega modula znotraj iste polipeptidne verige proteinskega origamija ta koeficient zmanjša. Efektivno se to kaže v večji verjetnosti neustrezno zvitih stanj (stanj, ki ne izkazujejo lastnosti poliedra). Da bi ugotovili, koliko identičnih modulov lahko uvedejo, da še ne pride do napačnega zvijanja, so pripravili tetraedrične proteinske origamije z dvema, uspela pa jim je tudi priprava s tremi identičnimi moduli od celokupno šestih, ki sestavljajo tetraedrično proteinsko kletko. Za uspešnost zvijanja je bilo ključnega pomena, ali so bili identični moduli sosednji. Rezultati so potrdili hipotezo, da kopičenje različnih motivov v polipeptidno verigo zmanjšuje verjetnost agregacije proteinov [4].
Zaključek
Raziskava med drugim prispeva k potrditvi, da zvijanje proteinskih molekul ne poteče usklajeno po korakih zgolj ene poti, hkrati pa tudi z vzpostavitvijo modela za napovedovanje zvijanja nakazuje, da teh poti ni neobvladljivo veliko. Slednja srednja pot mehanizma zvijanja proteinov, ki predvideva favoriziranje omejenega števila možnih vzporednih stopenj zvijanja, je bila predhodno nakazana že z drugimi študijami. Zaradi poudarjene modularnosti, ki je posledica specifične topološke zgradbe proteinskih origamijev, ti predstavljajo zanimiv in radikalno preprost model za nadaljnje študije zvijanja večjih modularnih proteinov. Za oceno razdalje med interagirajočima elementoma ovitih vijačnic so uporabili molekulsko dinamiko in eksperimentalne meritve kinetike zvijanja. Na podlagi rezultatov so razvili matematični model za napovedovanje verjetnosti zvijanja proteinskih origamijev z različnim številom osnovnih gradnikov.
Proteinski origamiji pa niso zanimivi zgolj z vidika preučevanja njihove dinamike zvijanja. Zaradi svoje značilne strukture in lastnosti (prilagodljivost, biorazgradljivost, sposobnost samosestavljanja, enostavnost za manipulacijo) imajo velik potencial v smeri razvoja biomedicinskih materialov, npr. kot dostavni sistemi za terapevtske učinkovine ali kot ogrodje za nekatera cepiva. Gre namreč za edini do sedaj poznani tip strukture proteinske kletke s središčno praznino, ki je sestavljen zgolj iz ene polipeptidne verige [10].
Viri
[1.] P. Gainza-Cirauqui, B. E. Correia: Computational protein design — the next generation tool to expand synthetic biology applications. Curr. Opin. Biotechnol. 2018, 52, str. 145–152.
[2.] S. Božič, T. Doles, H. Gradišar, R. Jerala: New Designed Protein Assemblies. Current Opinion in Chemical Biology. Elsevier Current Trends December 1, 2013, pp. 940–945.
[3.] A. Ljubetič, F. Lapenta, H. Gradišar, I. Drobnak, J. Aupič, Ž. Strmšek, D. Lainšček, I. Hafner-Bratkovič, A. Majerle, … R. Jerala: Design of coiled-coil protein-origami cages that self-assemble in vitro and in vivo. Nat. Biotechnol. 2017, 35(11), str. 1094–1101.
[4.] J. Aupič, Ž. Strmšek, F. Lapenta, D. Pahovnik, T. Pisanski, I. Drobnak, A. Ljubetič, R. Jerala: Designed folding pathway of modular coiled-coil-based proteins. Nat. Commun. 2021, 12(1), str. 1–12.
[5.] J. H. Han, S. Batey, A. A. Nickson, S. A. Teichmann, J. Clarke: The Folding and Evolution of Multidomain Proteins. Molecular Cell Biology. 2007, pp. 319–330.
[6.] C. P. Ponting, R. Mott, P. Bork, R. R. Copley: Novel protein domains and repeats in Drosophila melanogaster: Insights into structure, function, and evolution. Genome Res. 2001, 11(12), str. 1996–2008.
[7.] T. Tørring, N. V. Voigt, J. Nangreave, H. Yan, K. V. Gothelf: DNA origami: A quantum leap for self-assembly of complex structures. Chem. Soc. Rev. 2011, 40(12), str. 5636–5646.
[8.] I. Drobnak, A. Ljubetič, H. Gradišar, T. Pisanski, R. Jerala: Designed protein origami. Adv. Exp. Med. Biol. 2016, 940, str. 7–27.
[9.] Kemijski inštitut: Proteinski origami - nova generacija umetnih bionanostruktur. [pridobljeno 12. 4. 2021]. Dostopno na: https://www.ki.si/novice/single-prikaz/breaking/novica/proteinski-origami-nova-generacija-umetnih-bionanostruktur/
[10]. F. Lapenta, J. Aupič, Ž. Strmšek, R. Jerala: Coiled Coil Protein Origami: From Modular Design Principles towards Biotechnological Applications. Chemical Society Reviews. 2018, pp. 3530–3542.

{kind=link}
{kind=link}
{kind=link}