Odkritje in delovanje nukleaze Drosha: Difference between revisions
No edit summary |
AnaDolinar (talk | contribs) |
||
| (One intermediate revision by the same user not shown) | |||
| Line 8: | Line 8: | ||
== Odkritje == | == Odkritje == | ||
Drosha je bila odkrita pri Drosphilli (vinska mušica). Človeški homolog je bil sprva opisan kot protein, ki reagira s transkripcijskimi faktorji Sp1. Prvo raziskavo delovanja Droshe so opravili Stanley Crooke in sodelavci in so jo na podlagi rezultatov raziskave dodelili v družino ribonukleaz III. Dokazali so, da je Drosha jedrni protein. Opravili so eksperiment, kjer so z negativno regulacijo s protismiselnimi oligonukleotidi zmanjšali delovanje Droshe. Posledica zmanjšanega delovanja se je kazala v kopičenju intermediatov pri nastajanju pre-miRNA oz. kopičenju pri-miRNA. Tako so dokazali, da ima Drosha pomembno vlogo pri zorenju miRNA. Kasnejše raziskave so pokazale, da proces zorenja miRNA poteka v dveh stopnjah. V prvi stopnji encim Drosha v jedru prepozna substrat, začetni transkript oz. pri-miRNA, in ga cepi. Produkt cepitve je pre-miRNA, ki je dolga približno 70 nukleotidov. Pre-miRNA se nato prenese v citoplazmo, kjer poteče nadaljnja cepitev z encimom Dicer. Tako nastane zrela ~22 nukleotidov dolga molekula miRNA. | Drosha je bila odkrita pri ''Drosphilli'' (vinska mušica). Človeški homolog je bil sprva opisan kot protein, ki reagira s transkripcijskimi faktorji Sp1. Prvo raziskavo delovanja Droshe so opravili Stanley Crooke in sodelavci in so jo na podlagi rezultatov raziskave dodelili v družino ribonukleaz III. Dokazali so, da je Drosha jedrni protein. Opravili so eksperiment, kjer so z negativno regulacijo s protismiselnimi oligonukleotidi zmanjšali delovanje Droshe. Posledica zmanjšanega delovanja se je kazala v kopičenju intermediatov pri nastajanju pre-miRNA oz. kopičenju pri-miRNA. Tako so dokazali, da ima Drosha pomembno vlogo pri zorenju miRNA. Kasnejše raziskave so pokazale, da proces zorenja miRNA poteka v dveh stopnjah. V prvi stopnji encim Drosha v jedru prepozna substrat, začetni transkript oz. pri-miRNA, in ga cepi. Produkt cepitve je pre-miRNA, ki je dolga približno 70 nukleotidov. Pre-miRNA se nato prenese v citoplazmo, kjer poteče nadaljnja cepitev z encimom Dicer. Tako nastane zrela ~22 nukleotidov dolga molekula miRNA. | ||
== Struktura Droshe in Mirkoprocesor == | == Struktura Droshe in Mirkoprocesor == | ||
| Line 14: | Line 14: | ||
Encim Drosha sestavljata dve RNazni domeni, ki tvorita intramolekularni dimer, ki služi kot encimski katalitični center. Na 3`koncu verige pri-miRNA cepi domena RIIIDa, medtem ko domena RIIIDb cepi na 5`koncu verige, pri tem pa delujeta neodvisno druga od druge. Poleg RNaznih domen je na C- koncu še dsRBD domena, ki je sestavljena iz α heliksa (Ser1263-Thr1271), sledijo mu tri antiparalene β ploskve (Leu1283- Gly1314) in domena se konča z še enim α heliksom (Ile 1317-Lys-1331). Vse skupaj tvori αβββα strukturo. [http://www.rcsb.org/pdb/explore/images.do?structureId=2KHX Strukturo dsRBD domene] so razrešili s pomočjo NMR metode. Drosha ima poleg katalitičnih domen in dsRBD domene še N-terminalno domeno, ki predstavlja jedrni lokalizacijski signal. | Encim Drosha sestavljata dve RNazni domeni, ki tvorita intramolekularni dimer, ki služi kot encimski katalitični center. Na 3`koncu verige pri-miRNA cepi domena RIIIDa, medtem ko domena RIIIDb cepi na 5`koncu verige, pri tem pa delujeta neodvisno druga od druge. Poleg RNaznih domen je na C- koncu še dsRBD domena, ki je sestavljena iz α heliksa (Ser1263-Thr1271), sledijo mu tri antiparalene β ploskve (Leu1283- Gly1314) in domena se konča z še enim α heliksom (Ile 1317-Lys-1331). Vse skupaj tvori αβββα strukturo. [http://www.rcsb.org/pdb/explore/images.do?structureId=2KHX Strukturo dsRBD domene] so razrešili s pomočjo NMR metode. Drosha ima poleg katalitičnih domen in dsRBD domene še N-terminalno domeno, ki predstavlja jedrni lokalizacijski signal. | ||
Ker dsRBD domena ni dovolj za vezavo substrata na encim, drosha potrebuje partnerski protein, ki prepozna pri-miRNA. Imenuje se DGCR8 (DiGeorge syndrome critical region 8), znan kot Pasha pri Drosphilla melanogaster in C. elegans. Da je Drosha res odvisna od partnerskega proteina, dokazuje naslednji opravljen poskus. Skoraj povsem so izčrpali oz. zmanjšali koncentracijo DGCR8. Posledično je naraščala koncentracija pri-miRNA, medtem ko sta koncentraciji pre-miRNA in miRNA upadali. Tako so dokazali, da niti Drosha, niti DGCR8 sama nista bila aktivna, zato cepitev pri-miRNA ni potekla, medtem ko je kompleks obeh encimov postal aktiven. To je služilo kot dokaz, da je DGCR8 pomemben kofaktor encima Drosha. | Ker dsRBD domena ni dovolj za vezavo substrata na encim, drosha potrebuje partnerski protein, ki prepozna pri-miRNA. Imenuje se DGCR8 (DiGeorge syndrome critical region 8), znan kot Pasha pri ''Drosphilla melanogaster'' in ''C. elegans''. Da je Drosha res odvisna od partnerskega proteina, dokazuje naslednji opravljen poskus. Skoraj povsem so izčrpali oz. zmanjšali koncentracijo DGCR8. Posledično je naraščala koncentracija pri-miRNA, medtem ko sta koncentraciji pre-miRNA in miRNA upadali. Tako so dokazali, da niti Drosha, niti DGCR8 sama nista bila aktivna, zato cepitev pri-miRNA ni potekla, medtem ko je kompleks obeh encimov postal aktiven. To je služilo kot dokaz, da je DGCR8 pomemben kofaktor encima Drosha. | ||
DGCR8 vsebuje dve dsRBD domeni, ki prepoznata ssRNA segment na pri-miRNA in se zasidra na mesto med ssRNA (enovijačna RNA) in dsRNA (dvovijačna RNA) ter tako signalizira Droshi, da cepi ~11 bp stran od te povezave. V svoji osrednji regiji DGCR8 vsebuje tudi WW domeno (imenuje se tudi Rsp5/wwp), ki ima dva močno ohranjena triptofanska ostanka, ločena s približno 20 aminokislinami. Ta domena dobro reagira s prolini bogatimi regijami. Ker je Drosha bogata s prolini na N-koncu, lahko s kofaktorjem DGCR8 reagira preko domene WW. [http://c431376.r76.cf2.rackcdn.com/2282/fmicb-01-00108-HTML/image_m/fmicb-01-00108-g001.jpg Domene, ki tvorijo Drosho in DGCR8] | DGCR8 vsebuje dve dsRBD domeni, ki prepoznata ssRNA segment na pri-miRNA in se zasidra na mesto med ssRNA (enovijačna RNA) in dsRNA (dvovijačna RNA) ter tako signalizira Droshi, da cepi ~11 bp stran od te povezave. V svoji osrednji regiji DGCR8 vsebuje tudi WW domeno (imenuje se tudi Rsp5/wwp), ki ima dva močno ohranjena triptofanska ostanka, ločena s približno 20 aminokislinami. Ta domena dobro reagira s prolini bogatimi regijami. Ker je Drosha bogata s prolini na N-koncu, lahko s kofaktorjem DGCR8 reagira preko domene WW. [http://c431376.r76.cf2.rackcdn.com/2282/fmicb-01-00108-HTML/image_m/fmicb-01-00108-g001.jpg Domene, ki tvorijo Drosho in DGCR8] | ||
Latest revision as of 20:24, 4 April 2012
Drosha je proteinski kompleks, ki skupaj z DiGeorge syndrome critical region 8 (DGCR8) cepi primarne lasnične miRNA (pri-miRNA) molekule v prekurzorske miRNA (pre-miRNA). Je endonukleaza, ki cepi na obeh verigah RNA heliksa. Cepi samo dvoverižne RNA, enoverižnih ne cepi. Encim Drosha sestavljajo dve RNazni domeni, ki tvorita encimski katalitični center. Poleg RNaznih domen je na C- koncu še dsRBD domena.
Osnove
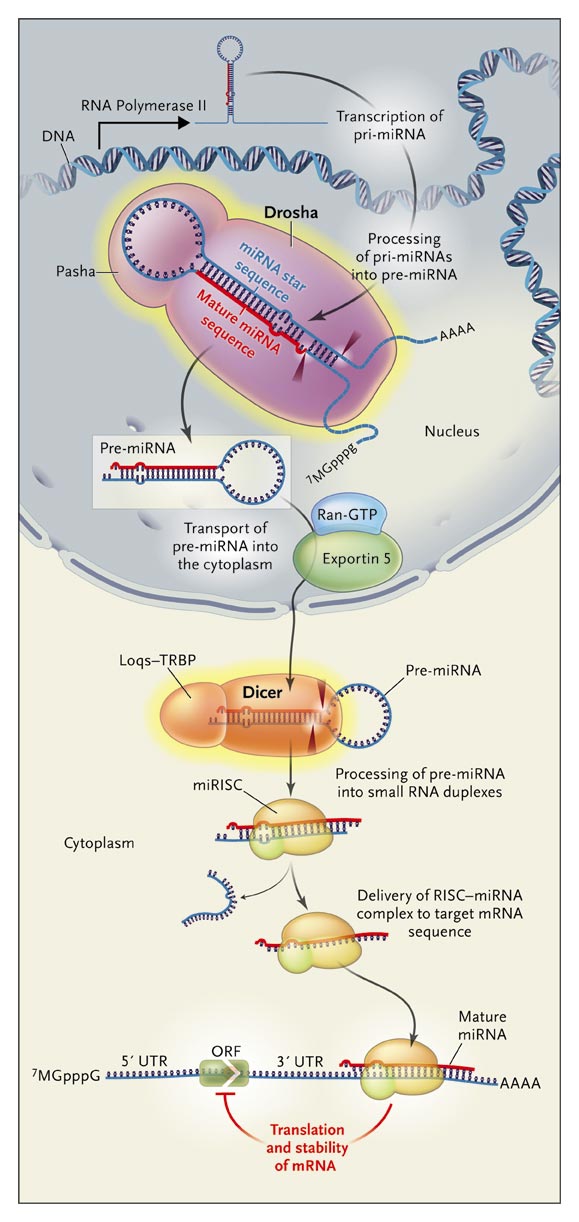
MikroRNA (miRNA) je enovijačna RNA (ssRNA), dolga približno 22 nukleotidov. Je regulatorna molekula, ki nadzoruje različne celične funkcije v evkariontih, kot so diferenciacija, razvoj in protivirusna obramba. Proces zorenja miRNA poteka v jedru in v citoplazmi in sicer v dveh stopnjah. Začne se v jedru, kjer najprej poteče transkripcija z encimom RNA polimeraza II. Začetni transkript, ki nastane pri transkripciji (pri-miRNA) cepi encim Drosha na približno 70 nukleotidov dolgo prekurzorsko miRNA (pre-miRNA). Nastalo pre-miRNA encim Eksportin 5 prenese iz jedra v citoplazmo, kjer jo encim Dicer dalje cepi na miRNA. Drosha je specifična endonukleaza, ki cepi dvovijačno RNA (dsRNA) tako, da reže na vsaki od verig dvovijačnice oz. heliksa. Velika je približno 160 kDa. Spada v družino ribonukleaz III (RNaz III), ki jih najdemo tako pri evkariontih kot pri prokariontih. Ribonukleaze III se delijo na 3 razrede glede na razporeditev domen. Drosho uvrščamo v II. razred ribonukleaz III, saj vsebuje dve katalitični domeni in C-terminalno domeno dsRBD oz. dsRNA-binding domain. Poleg tega ima Drosha še eno domeno, in sicer N-terminalno domeno, katere vloga naj bi bila signalizacijska sekvenca, ki služi kot signal transporta Droshe v jedro. Za razliko od razreda II so encimi I. razreda preprostejši. Imajo le eno katalitično domeno in domeno dsRBD. Razred III, v katerega uvrščamo encim Dicer, vsebujejo helikazno domeno, PAZ domeno in DUF283 domeno, ne glede na nukleazno in dsRBD domeno. Človeški genom kodira le dve ribonukleazi III – Dicer in Drosho. Homologi Droshe so prisotni le pri živalih.
Odkritje
Drosha je bila odkrita pri Drosphilli (vinska mušica). Človeški homolog je bil sprva opisan kot protein, ki reagira s transkripcijskimi faktorji Sp1. Prvo raziskavo delovanja Droshe so opravili Stanley Crooke in sodelavci in so jo na podlagi rezultatov raziskave dodelili v družino ribonukleaz III. Dokazali so, da je Drosha jedrni protein. Opravili so eksperiment, kjer so z negativno regulacijo s protismiselnimi oligonukleotidi zmanjšali delovanje Droshe. Posledica zmanjšanega delovanja se je kazala v kopičenju intermediatov pri nastajanju pre-miRNA oz. kopičenju pri-miRNA. Tako so dokazali, da ima Drosha pomembno vlogo pri zorenju miRNA. Kasnejše raziskave so pokazale, da proces zorenja miRNA poteka v dveh stopnjah. V prvi stopnji encim Drosha v jedru prepozna substrat, začetni transkript oz. pri-miRNA, in ga cepi. Produkt cepitve je pre-miRNA, ki je dolga približno 70 nukleotidov. Pre-miRNA se nato prenese v citoplazmo, kjer poteče nadaljnja cepitev z encimom Dicer. Tako nastane zrela ~22 nukleotidov dolga molekula miRNA.
Struktura Droshe in Mirkoprocesor
Encim Drosha sestavljata dve RNazni domeni, ki tvorita intramolekularni dimer, ki služi kot encimski katalitični center. Na 3`koncu verige pri-miRNA cepi domena RIIIDa, medtem ko domena RIIIDb cepi na 5`koncu verige, pri tem pa delujeta neodvisno druga od druge. Poleg RNaznih domen je na C- koncu še dsRBD domena, ki je sestavljena iz α heliksa (Ser1263-Thr1271), sledijo mu tri antiparalene β ploskve (Leu1283- Gly1314) in domena se konča z še enim α heliksom (Ile 1317-Lys-1331). Vse skupaj tvori αβββα strukturo. Strukturo dsRBD domene so razrešili s pomočjo NMR metode. Drosha ima poleg katalitičnih domen in dsRBD domene še N-terminalno domeno, ki predstavlja jedrni lokalizacijski signal.
Ker dsRBD domena ni dovolj za vezavo substrata na encim, drosha potrebuje partnerski protein, ki prepozna pri-miRNA. Imenuje se DGCR8 (DiGeorge syndrome critical region 8), znan kot Pasha pri Drosphilla melanogaster in C. elegans. Da je Drosha res odvisna od partnerskega proteina, dokazuje naslednji opravljen poskus. Skoraj povsem so izčrpali oz. zmanjšali koncentracijo DGCR8. Posledično je naraščala koncentracija pri-miRNA, medtem ko sta koncentraciji pre-miRNA in miRNA upadali. Tako so dokazali, da niti Drosha, niti DGCR8 sama nista bila aktivna, zato cepitev pri-miRNA ni potekla, medtem ko je kompleks obeh encimov postal aktiven. To je služilo kot dokaz, da je DGCR8 pomemben kofaktor encima Drosha. DGCR8 vsebuje dve dsRBD domeni, ki prepoznata ssRNA segment na pri-miRNA in se zasidra na mesto med ssRNA (enovijačna RNA) in dsRNA (dvovijačna RNA) ter tako signalizira Droshi, da cepi ~11 bp stran od te povezave. V svoji osrednji regiji DGCR8 vsebuje tudi WW domeno (imenuje se tudi Rsp5/wwp), ki ima dva močno ohranjena triptofanska ostanka, ločena s približno 20 aminokislinami. Ta domena dobro reagira s prolini bogatimi regijami. Ker je Drosha bogata s prolini na N-koncu, lahko s kofaktorjem DGCR8 reagira preko domene WW. Domene, ki tvorijo Drosho in DGCR8
Drosha in DGCR8 se povežeta med seboj in tako tvorita funkcionalni kompleks, znan kot Mikroprocesor, 500- 650 kDa velik encimski kompleks. Mikroprocesor katalizira cepitev dolgih primarnih mi-RNA transkriptov (pri-mi RNA) v kratke lasnice (pre-mi RNA), dolge približno 70 nukleotidov. Ključno vpliva na specifičnost miRNA, saj določi kje bo deloval naslednji RNAzni encim Dicer. Vse pri-miRNA vsebujejo terminalno zanko, so dvovijačne molekule in vsebujejo sekvenco, ki je enovijačna. In ravno te značilnosti omogočajo mikroprocesorju prepoznavanje pravega mesta cepitve. Raziskave so pokazale, da lahko Mirkoprocesor vsebuje več kopij proteina Droshe ali DGCR8, ali drugih proteinov povezanih z Drosho in DGCR8.
Mikroprocesor ima večjo afiniteto in specifičnost do substrata, kot bi jo imela posamezna encima Drosha in DGCR8, vezana na pri-miRNA. 5` in 3` konec pri-miRNA sta osnovna segmenta začetnega transkripta RNA, nadaljujeta se v spodnjo dvovijačno RNA, ki poteka od osnovnih segmentov do mesta cepitve, od mesta cepitve pa do terminalne oz. končne zanke pa poteka zgornja dvovijačna RNA. dsRBD domena proteina DGCR8 lahko prepozna oba dela dvovijačne RNA, spodnjega in zgornjega. Domena dsRBD encima Drosha pa se lahko veže na centralno mesto blizu mesta cepitve pri-miRNA. Drosha deluje kot katalitična podenota, medtem ko je DGCR8 (Pasha) ključen za prepoznavanje substrata (pri-mi RNA). Dve podenoti encima Drosha interagirata med seboj in ustvarita intramolekularni dimer, kjer vsaka od katalitičnih enot cepi eno od dsRNA. RIIIDb cepi na 5` strani lasnice in RIIDa cepi na 3` strani. To povzroči, da se pojavita na 3` koncu dva prosta nukleotida, ki ˝štrlita˝ ven- štrleča konca. Ta dva prosta nukleotida sta zelo pomembna, saj ju encim Eksportin 5 prepozna in tako omogoči prenos pre-mi RNA iz jedra v citoplazmo.
Regulacija
Mikroprocesor je pri biogenezi miRNA prvi člen regulacije. Regulacija poteka preko uravnavanja mikroprocesorjeve aktivnosti. Drosha in DGCR8 sta regulirana tako transkripcijsko kot postranskripcijsko. Če se koncentracije Droshe in DGCR8 zvišane v celici, Mikroprocesor cepi DGCR8 mRNA. To povzroči zmanjšano sintezo DGCR8 in s tem destabilizacijo proteina Droshe. Kar privede do inaktivacije Mikroprocesorja. Mikroprocesor ima lastnosti avtoregulacije. Proteini, ki vplivajo na delovanje- regulacijo mikroprocesorja so t.i. DEAD box proteini. To sta predvsem proteina p68 in p72. Sta od ATP odvisna helikazna proteina. Poleg beljakovinskega kompleksa spliceosoma sta pomembna pri izrezovanju intronov in opravljata helikazno funkcijo. Ta dva proteina vplivata na mnoge druge proteine in transkripcijske faktorje, kot so: Smads, p53 in receptorji za estrogen. Smads so transkripcijski faktorji, ki so pozitivni regulatorji Droshe. So signalne molekule družine TGFβ. Smads regulirajo gensko ekspresijo že v samem začetku, tako, da vplivajo na transkripcijo DNA ali pa postranskripcijsko vplivajo na zorenje miRNA tako, da se v jedru vežejo na Drosho in p68 ter omogočijo Droshi vezavo na pravo mesto za cepitev. P53 je tudi pozitivni regulator nastanka miRNA. p53 deluje tako, da se poveže z helikazo p68 in olajša inerakcijo Drosha-pri-miRNA. Receptor za estrogen (ERα) je naegativni regulator in deluje tako, da se poveže z estradiolom in z p68 ali p72 ter povzroči, da se Drosha odcepi od pri-mi RNA in s tem prepreči nadaljno zorenje miRNA. (http://jb.oxfordjournals.org/content/148/4/381/F2.expansion.html)
Literatura
- Carmichael, G.G. RNA silencing: Methods and protocols. New Yersey, Humana Press, 2005
- Dusebery D.N.B., Hata A. Mechanisms of control of micro RNA biogenesis. The journal of biochemistry.2010, 148,4:381-392
- Yeom et. al. Caracterization of DGCR8/Pasha, the essential cofactor for Drosha in primary miRNA processing. Nucelic Acid Research, 2006, vol. 34, no. 16, 4622-4629
- Mueller et. al. Solution structure of the Drosha double-stranded RNA-binding domain. Silence, 2010, 1-5
- Zeng, Y., Ruy, Y. in Cullen, B.R. Recognition and cleavage of primary microRNA precursor by the nuclear processing enzyme Drosha. The EMBO journal, 2005, vol. 24, no. 1, 138-148

{kind=link}
{kind=link}