Parvovirusi in sorodni ssDNA-virusi
ssDNA virusi
Virusi, ki vsebujejo enoverižno DNA so zelo razširjeni v okolju. Prisotni so v morski vodi, v sladki vodi, v sedimentih in v ekstremnih okoljih. Enoverižen genom, ki ga vsebujejo parvovirusi je negativno nabit. Virusi ssDNA so razvili različne mehanizme za napad na celične genome, ki se pretežno razlikujejo med virusi, ki okužijo bakterije in evkarionte.
Parvovirusi
Virusi iz družine Parvoviridae so najenostavnejši živalski in človeški DNA virusi. Latinska beseda 'parvus' razlaga njihovo najbolj pomembno lastnost, da so majhni. Ta lastnost veliko vpliva na njihovo pomnoževanje, ki je v velikem odvisno od celičnih mehanizmov gostitelja. Poznamo več kot 50 tipov parvovirusov, ki so razdeljeni v dve poddružini: Parvoviridae, ki okužijo vretenčarje in Densoviridae, ki so karakteristični za okužbo členonožcev in zuželk. Poddružina Parvoviridae je nadalje razdeljena na pet rodov: amidovirusi, bokavirusi, dependovirusi, eritrovirusi in humani parvovirusi. Večina parvovirusov, kot so eritrovirusi, se podvojuje avtonomno, pri nekaterih pa virusno pomnoževanje ni možno brez sočasne prisotnosti virusov pomočnikov. Primer virusov, odvisnih od virusov pomočnikov so dependovirusi oz. adeno povezani virusi (AAV).
Struktura parvovirusov
Parvovirusi so majni, ikozaedrični virusi brez ovojnice, s premerom okrog 26 nm. Genom parvovirusa predstavlja linearna enovijačna DNA, velikosti približno 3 000 -5 000 baz. Svojo nukleinsko kislino virusi ščitijo s posebnim membranskim plaščom, kapsidom. Ta embalaža mora biti dovolj odporna, da zaščiti genom pred zunanjimi vplivi, ampak istočasno ustrezno labilna, da se lahko vsebina sprosti, ko pride v primerno okolje. Strukture kapsidov za več parvovirusov so bile določene z rentgensko kristalografijo in krioelektronsko mikroskopijo. Virusni proteini, VP1, VP2 in VP3, ki sestavljajo kapsido vsebujejo ohranjene in varijabilne regije (VR). Ohranjene regije so β-sodček in α-vijačni del. Biokemijske in strukturne raziskave kažejo da so ostanki v VR regijah zelo pomembni pri vezavi virusa na virusne receptorje in pri celotni virusni okužbi. Za površino kapsidov so značilne številne depresije in izbokline, ki pomagajo pri vezavi virusa na gostiteljsko celico. Proteinski plašč in nukleinska kislina skupa činijo nukleokapsid. Nekateri parvorisusi so s časom formirali dodatno zaščito, v obliki dodatke lipidne opne.
Vezava na gostiteljsko celico
Različni virusi so razvili različne poti za okužbo gostiteljskih celic. Okužba celic je običajno večstopenjski proces, katerega je prvi korak vezava virusa na celično površino. Pritrditev virus na gostiteljsko celico je posredovana preko virusnih površinskih komponent (površine kapside), ki se vežejo na specifične molekule na površini gostiteljsko celice, kot so membranski proteini, lipidi ali ogljikovi hidrati. Za identifikacijo kapsidnih aminokislinskih ostankov, ki prepoznajo receptorje na površini celične membrane so uporabljene analize vezave celic, transdukcijske analize in strukturni procesi. Analize so pokazale da obstajajo mesta v VR regijah kapside AAV, ki služijo kot vezavne regije, neodvisno od prepoznanega glikana. Virusi prepoznajo glikane na površini kot primarne receptorje. Adeno vezani virusi, oz. dependovirusi (AAV2, AAV3, AAV6 in AAV13) se vežejo na heparan sulfat preoteoglikan (HSPG). AAV1, AAV4 in AAV5 vežejo sialno kislino med okužbo celic, AAV9 pa N-vezavno galaktozo (GAL). Glikan glikoforin je karakterističen za avtonomni parvorisus, bokavirus. B19 eritrovirus pa se veže na glikosfingolipidni eritroitni P antigen (glubosid Gb4) . Vezavna mesta za glikane se prekrivajo z učinkovitostjo transdukcije in patogenosti virusov.
Vstop virusa v okuženo celico
Celica nalaga veliko ovir za vnos virusa. Kljub temu, virusi izkoriščajo temeljne celične procese da vstopijo i sprostijo svoj genski tovor. Vsi parvovirusi potrebujejo receptorsko posredovano endocitozo za okužbo celic. Po vezavi virusa na receptorje se virus obvije s klatrini, in na ta način formira klatrinski vezikel. Temu sledi vstop vezikla v celico, pot do zgodnjega endosoma in poznega endosoma. Klatrinski plašč se odstrani in se virus sprosti v citosol. Ko so v notranjosti celice, kapside se spremenijo, pri čemer izpostavijo N končna dela kapsidnih proteina VP1 in VP2 in 3' konec virusnega genoma. Na kapside vpliva protealitična aktivnost proteasomov, in se takrat verjetno formira tretji kapsidni protein VP3. Izpostavljen N-končni del VP1 kapsidnega proteina bo služil kot jedrni lokalizacijski signal. Virusi, ki se podvojujejo v jedru, potrebujejo mehanizme za prenos kapside in genoma do jedrnih por. V citoplazmi je prisoten citoskelet, čigave komponente virus izkorišča kot tire do jedra. Številni eksperimenti so pokazali da CPV (cannine parvovirus) kapside znotraj citoplazme potrebovali 3 do 6 ur da bi vstopile v jedro. Prevoz do jedra so preprečili z depolimerizacijo mikrotubulov in blokiranjem motoričnega proteina mikrotubulov, dineina, ki prenaša tovor proti minus koncu mikrotubula, ki je v neposredni bližini jedra. S tem so ugotovili da so mikrotubuli in dineini ključni za prenos kapside virusa do jedra. Virus ssDNA je dovolj majhen da vstopi v jedro preko jedrsko pornog kompleksa (NPC). Večji, dsDNA virusi, pa potrebujejo importin za vstop v jedro.
Vstop v jedro
Transport kapsida je omogočen s pomočjo ATP in Ran-1 proteina. Tisti kapsidi, ki pridejo v celico imajo 20 do 30 nukleotidov izpostavljenih v obliki špange, kateri nastajajo zaradi končnih obrnjenih ponovitev. Končne obrnjene ponovitve, ali TIR (terminal inverted repeats), so sestavljene iz palindromskih sekvencah ki so večinoma enake s obe strani genoma. Ta izpostavljen del DNA služi kod začetni oligonukleotid s pomočjo katerega popravljalni encimi okužene celice pretvorijo ssDNA virusa v dsDNA, dsDNA virusa je potem pripravljena za transkripcijo in translacijo s pomočjo celičnih proteinov. TIR omogoča tudi istočasno sproščanje virusnega DNA brez razstavljanja kapsida.
Ekspresija genov
Genom je podeljen v dva dela, desna stran kodira strukturne proteine, leva pa regulatorne. Število promotorjev se razlikuje od vrste do vrste (od 1 do 3). Zaradi izredno majhne dolžine genoma, če je več promotorjev, oni se prekrivajo. Genom pravovirusov kodira regulatorne in strukturne proteine s prekrivajočim se sekvencami, kar je omogočeno s pomočjo alternativnega izrezovanja intronov oz. spajanja eksonov. Izražanje genov pa regulirajo proteini rep (naziv so dobili ker mutacija gena za izražanje tega proteina tudi onemogoča replikacijo virusne DNA) pri AAV in NS (non structural) pri samostojnih virusih. Večina regulatornih proteinov se ne vežejo direktno na DNA, ampak aktivirajo ali inhibirajo proteine v inficirani celici, kar povzroča manjše ali večje izražanje določenih genov na virusni DNA. Poleg tega, strukturni proteini so tudi pokazali da imajo vlogo v regulaciji gena.
AAV imajo značilno karakteristiko da v večini zdravih celicah nimajo sposobnost normalne aktivnosti translacije in transkripcije dokler celica ne pride v toksično okolje. Toksično okolje so lahko ekstremne temperature, UV radiacija ali kemijski kancerogeni, ampak najbolj učinkovit aktivator AAV so adenovirusi in herpsvirusi. Geni tistih virusov imajo več vlog za AAV. Ampak, vpliv tih genov ni nujen, AAV bo stabilno bival v celici dokler je celica zdrava, ko se pojavijo znaki poškodovanja celice replikacija AAV se aktivira in virus išče novega gostitelja. Pri translaciji AAV gena za VP2 lahko pride do leaky scanning (prepuščajočeg skeniranja) pri čemer se translacija ne začne na slabem začetnem ACG kodonu da bi potem nastal VP2, ampak na močnem AUG in nastane VP3.
Replikacija
ssDNA virusi uporabljajo tako imenovan “rolling hairpin” mehanizem za svojo replikacijo ( https://viralzone.expasy.org/resources/ssstrand_displacement.jpg ). Ta mehanizem ima dve specifične karakteristike. Prva je da se TIR lahko uporablja kot začetni oligonukleotid za začetek replikacije in druga je da ni zaostajajoče verige pri replikaciji. Replikacija je aktivna samo med S fazo celičnega cikla za samostojne viruse in v toksičnih pogojih za AAV. Aktivira je virusna endonukleaza rep pri AAV ali NS1 pri samostojnih virusih. Rep naredi zasek med kodirajočo regijo in regijo katera ustvarja “hairpin” na 5’ koncu. Ta zasek omogoča pomik verige in začetek translacije. Ko se en cikel replikacije konča virusi kateri imajo enake, ali podobne končne obrnjene ponovitve proizvajajo enako število ssDNA+ in ssDNA-. Tisti virusi ki nimajo podobne končne obrnjene ponovitve pa proizvajajo večinoma ssDNA- verige, dokler se ssDNA+ reciklira za nov cikel.
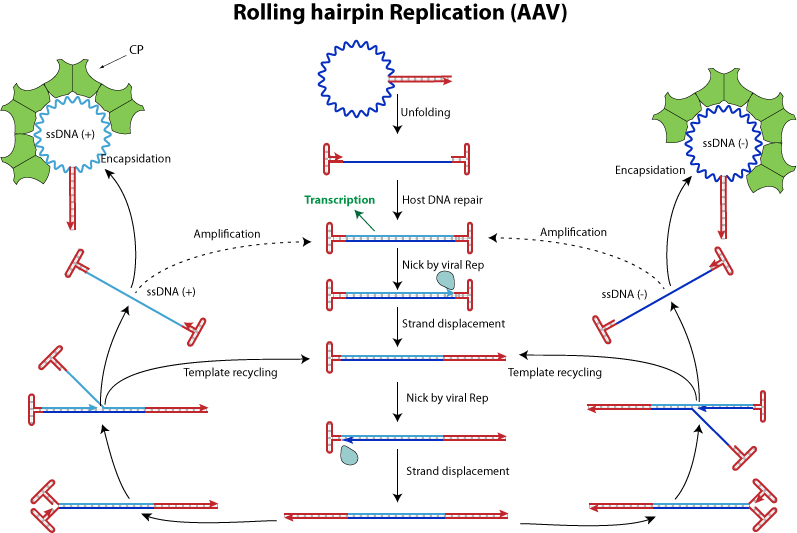{kind=link}
Integracija AAV v gostiteljsko DNA
AAV ima preferenco za določeno regijo na ljudskem kromosomu, AAVS1, ampak se tudi veže na druge regije ki imajo zavezujoči motiv rep protein vključen v integracijo virusa. Ta motiv se tudi nahaja na delu v obliki špange AAV virusa in je sestavljen iz Rep-zavezujočeg mesta(RBS, rep-binding site) in končno resolucijsko mesto (TRS, terminal resolution site). Rep protein ima sposobnost da veže istočasno TRS regijo AAV DNA in DNA gostitelja pri čemer privezuje oba genoma.
Rakotvornost
Za razliko od ostalih DNA virusov kateri lahko vzpostavijo stabilen odnos z gostiteljsko celico, parvovirusi niso pokazali rakotvorna dejstva, nasprotno, pokazili so tudi da lahko imajo preventivni efekt. Ta značilnost, predvsem dependovirusov, je omogočena zaradi toksične sredine v kateri se treba nahajati virus da bi prišlo do replikacije. To sredino lahko najdemo predvsem v transformiranih in kancerogenih celicah. Reprodukcija AAV bo imela negativen vpliv na te celice in s tem bo prispevala do upočasnitve tumorigeneze.
Viri
- Berns, K. I. (1990) ‘Parvovirus Replication’, MICROBIOLOGICAL REVIEWS, 54(3), pp. 316–329. Available at: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC372780/pdf/microrev00038-0114.pdf (Accessed: 12 April 2018).
- Grove, J. and Marsh, M. (2011) ‘The cell biology of receptor-mediated virus entry’, Journal of Cell Biology, 195(7), pp. 1071–1082. doi: 10.1083/jcb.201108131.
- Huang, L. Y., Halder, S. and Agbandje-Mckenna, M. (2014) ‘Parvovirus glycan interactions’, Current Opinion in Virology. Elsevier B.V., 7(1), pp. 108–118. doi: 10.1016/j.coviro.2014.05.007.
- Krupovic, M. and Forterre, P. (2015) ‘Single-stranded DNA viruses employ a variety of mechanisms for integration into host genomes’, Annals of the New York Academy of Sciences, 1341(1), pp. 41–53. doi: 10.1111/nyas.12675.
- Vihinen-Ranta, M., Suikkanen, S. and Parrish, C. R. (2004) ‘Pathways of cell infection by parvoviruses and adeno-associated viruses.’, Journal of virology. American Society for Microbiology, 78(13), pp. 6709–14. doi: 10.1128/JVI.78.13.6709-6714.2004.