Biokemijska struktura telomerov
Uvod
Za ohranjanje koncev linearnih kromosomov se morajo celice spopasti z dvema osrednjima biološkima problemoma: poskrbeti morajo za njihovo zaščito in podvajanje.
Konci evkariontskih kromosomov se morajo torej razlikovati od koncev, ki nastanejo zaradi poškodb DNA in jih je potrebno popraviti. Če se med seboj ne bi razlikovali, bi lahko prišlo do zlitja kromosomov, kar bi bilo za celico škodljivo. Z zaščito koncev kromosomov se prepreči ravno takšne reakcije.
Drugi problem predstavlja nepopolno delovanje DNA polimeraze, saj se konci kromosomov pri vsakem podvajanju skrajšajo. DNA polimeraza hčerinsko verigo sintetizira od 5ʹ proti 3ʹ koncu in ni zmožna zapolniti vrzeli, ki nastane na skrajnem koncu zaostajajoče matrične verige (Slika 1). Evkariontske celice so ta problema rešile z oblikovanjem posebnih struktur, imenovanih telomeri.
Struktura telomernih regij DNA
Telomeri so s številnimi proteini povezana področja ponavljajočih se nukleotidnih zaporedij na koncih linearnih kromosomov v jedrih evkariontskih celic. Njihova glavna vloga je zaščita koncev kromosomov pred razgradnjo in fuzijo s konci drugih kromosomov. Telomerna DNA je sestavljena iz kratkih (vendar pri različnih organizmih različno dolgih) z gvaninom bogatih ponavljajočih se zaporedij. V sesalskih celicah te velike nukleoproteinske komplekse gradijo dvovijačne tandemske šestnukleotidne ponovitve zaporedja TTAGGG, ponekod najdemo tudi variacije v obliki TTGGGG, TCAGGG in TGAGGG. Na skrajnem koncu jim sledi enoverižen 3ʹ razmeroma homogen, pretežno gvaninski previs. Posamezen telomer lahko vsebuje le nekaj tisoč ali pa celo več kot sto tisoč baznih parov ponavljajočega se zaporedja, pri ljudeh deset do dvajset tisoč baznih parov.
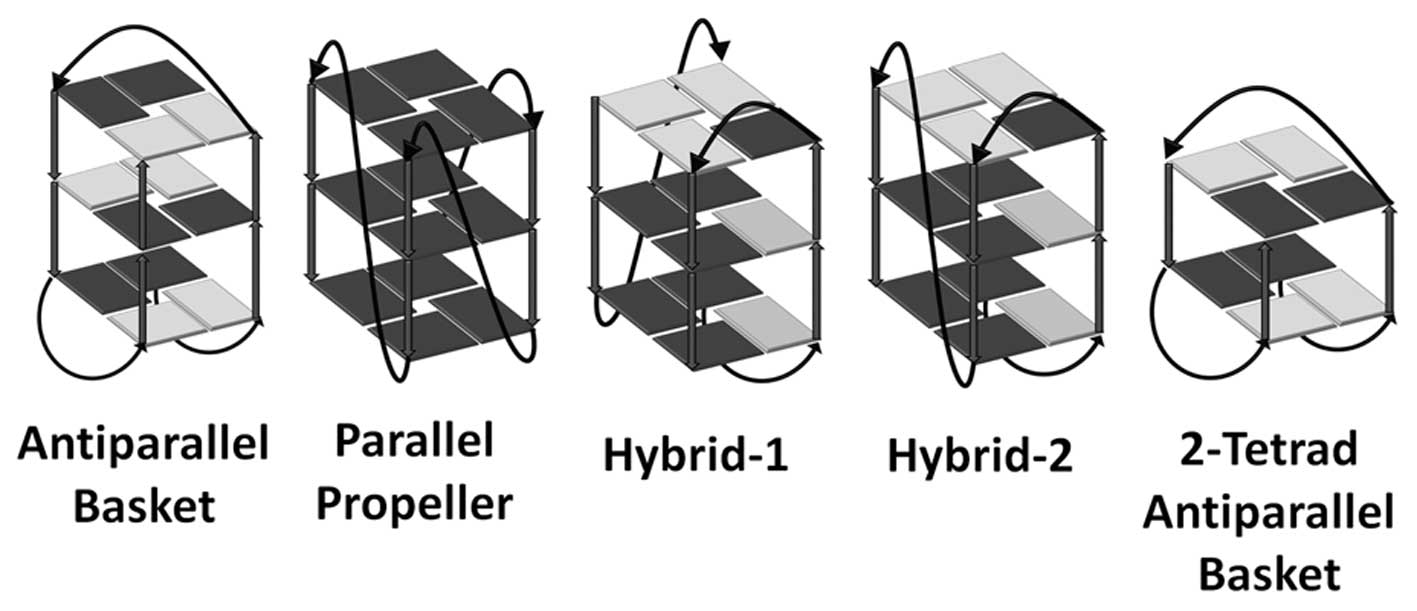
Večina te DNA naj bi tvorila nukleosome (en nukleosom tvori približno 146 baznih parov, ki so 1,67-krat oviti okoli histonskega oktamera), v bližini skrajnega konca pa se pojavljajo tako imenovani G-kvadrupleksi. Gre za večverižne strukture, ki jih skupaj držijo kvadratne ravnine štirih gvaninov (G-kvarteti), povezani s Hoogsteenovimi vodikovimi vezmi. Zavzemajo lahko različne konformacije. Nekatere so prikazane na sliki 2. Struktura takih tetrad je običajno še dodatno stabilizirana s kalijevim kationom, ki zavzame položaj med paroma tetrad. Tvorba G-kvadrupleksov lahko predstavlja problem pri podvajanju DNA in pri podaljševanju telomer s telomerazo.
{kind=link}
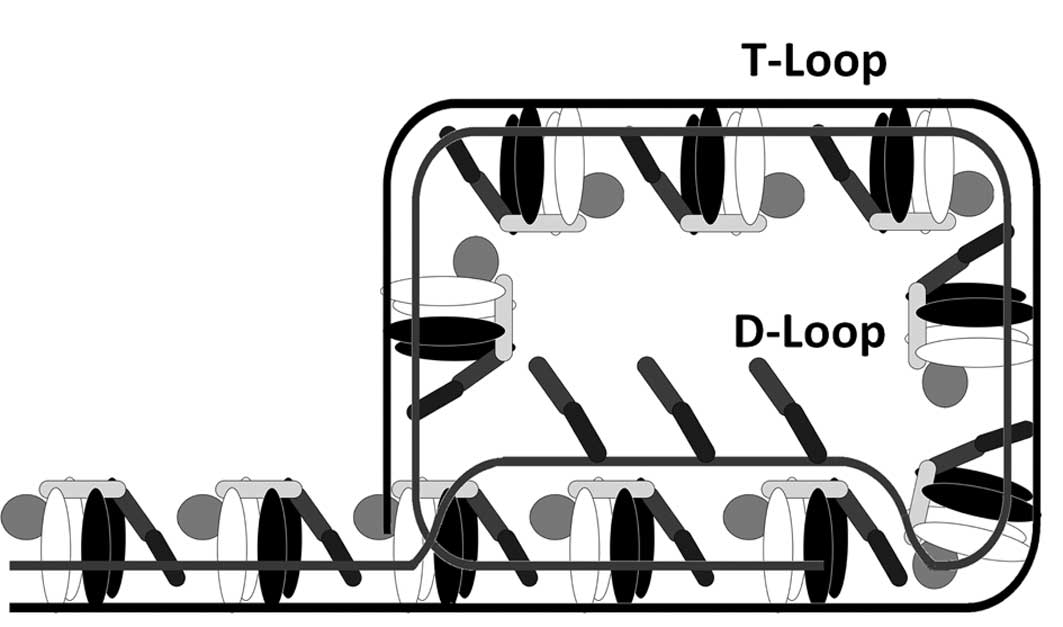
Telomerna DNA tvori velike T-zanke (telomere loop), ki zaključijo konce kromosomov. Pri tem se enoverižni telomerni konec s pomočjo proteinov uviha in napade dvoverižno DNA. Nastane triverižna struktura, tako imenovana D-zanka (displacement loop) (Slika 3). Vsaka gvaninska baza z enoverižnega 3ʹ konca služi hkrati kot donor in kot akceptor elektronov za tvorbo vodikovih vezi. Tako nastala kapi podobna struktura telomere varuje pred tem, da bi jih endonukleaze prepoznale kot poškodovano dvoverižno DNA in sprožile proces razgradnje.
{kind=link}
Struktura telomerov naj bi bila med celičnim ciklom dokaj dinamična, predvsem med fazo S, ko so telomeri podvrženi replikaciji.
Vezavni proteini telomerov
Telomerni DNA se v procesu pokrivanja pridružijo specifični vezavni in adapterski proteini, ki ohranjajo zaključeno strukturo in preprečujejo iniciacijo mehanizmov odgovora na poškodbe dednega materiala (DDR). Kljub ohranjenosti funkcij telomerov pri različnih organizmih sta zgradba in aminokislinsko zaporedje teh proteinov presenetljivo zelo raznolika, kar kaže na njihovo hitro spreminjanje tekom evolucije. Kljub temu pa jih glede na mesto in tip vezave lahko razvrstimo v tri večje skupine:
- dsDNA (dvoverižna DNA) vezavni proteini,
- ssDNA (enoverižna DNA) vezavni proteini in
- adapterski proteini.
Pri sesalcih se sestavijo v multiproteinski kompleks, imenovan šelterin oziroma telosom (Slika 4). Sestavlja ga šest komponent: TRF1 in TRF2, TIN2, RAP1 ter POT1 in TPP1.
TRF1, ki je bil odkrit prvi, služi kot negativni regulator podaljševanja dolžine telomerov s telomerazo. Poveča tudi učinkovitost podvojevanja TTAGGG ponovitev s preprečevanjem zastajanja replikacijskih vilic in sodeluje pri organizaciji telomernega kromatina.
TRF2 omogoči nastanek D-zanke z vključitvijo 3' previsa v dsDNA in tako zaključi T-zanko. Preko interakcije z raznimi proteinskimi faktorji je vključen tudi v zaščito telomernih koncev, saj njegova odsotnost vodi do fuzije telomerov in ATM ali p53 inducirane apoptoze.
POT1 z vezavo stabilizira 3' konec ssDNA in ga ščiti pred mehanizmi odziva na poškodbe DNA. Udeležen je tudi pri kontroli telomeraznega podaljševanja DNA, saj lahko v kompleksu s TPP1 deluje bodisi kot njen inhibitor bodisi kot procesivnostni faktor.
Proteini TIN2, RAP1 in TPP1 spadajo v zadnjo skupino, saj s prej omenjenimi tvorijo različne komplekse. TIN2 z direktno vezavo na TRF1, TRF2 in TPP1/POT1 heterodimer deluje kot centralna komponenta telosoma, saj v njegovi odsotnosti ne more priti do sestavljanja tega kompleksa. RAP1 služi kot adapterski protein za TRF2, saj z vezavo poveča afiniteto TRF2 do dsDNA in preprečuje nehomologno povezovanje koncev (NHEJ).
Šelterin ščiti sesalčje telomere pred popravljalnimi mehanizmi DDR, ohranja strukturo v obliki T-zanke in s preprečevanjem telomeraznega podaljševanja omejuje njihovo dolžino.
Model treh stanj telomerov skozi celični cikel normalne in rakave celice
Telomeri lahko skozi celični cikel tvorijo tri različne oblike: zaprto (closed-state), odprto (intermediate-state) in nepokrito (uncapped-state) (Slika 5).
Zaščiteni telomeri so v fazi G1 organizirani v zaprto obliko, tekom replikacije pa se pasivno ali aktivno razprejo, preidejo v odprto obliko. Pasivni način razpiranja omogoči vzdolž verige potujoča DNA polimeraza, aktivni pa RTEL helikaza. V S fazi celičnega cikla se telomerni konci podvojijo, pri čemer sodeluje protein TRF1. Po končani replikaciji šelterinska proteina TRF2 in POT1 skupaj s kompleksom Apollo in CST dokončno izoblikujejo 3' konec. V G2 fazi nato sledi pakiranje telomerov v zaščiteno, zaprto obliko, za kar so najverjetneje odgovorni TRF2 in drugi proteini.
Kot povsod drugod tudi tu lahko pride do zapletov. Zgodi se, da se med fazo G2 odprta oblika telomera zaradi pomanjkanja TRF2 proteina ne more preoblikovati v zaprto obliko. To je navadno značilno le za eno izmed dveh sestrskih kromatid kromosoma. Sprememba v kromosomu se kljub temu prenese v nov celični cikel, hkrati pa sproži popravljalni mehanizem (DDR). Če je število takih sprememb (telomeri odprtih oblik) večje od pet, se zaradi delovanja tumorskega zaviralnega gena p53 rast celice in celični cikel ustavita. Za rakave in modificirane celice, ki nimajo p53 zaviralne poti, pa je značilno, da se delijo, čeprav je na koncih njihovih kromosomov več kot pet telomerov odprtih oblik.
Do druge nezgode pa lahko pride v fazi M, ko naj bi se zaradi ionizirajočega sevanja preko proteina Aurora B aktivirala ATM kinaza, ki povzroči konformacijo nepoškodovane, zaprte oblike telomere v odprto. Pri tem se velik delež TRF2 proteinov odcepi od telomerov, preostali pa skrbijo, da se odprte telomere posameznih kromosomov med seboj ne zlepijo (Slika 6).
Pri normalnih celicah se po številnih delitvah telomere skrajšajo tako močno, da preidejo v nepokrito obliko. Pri tej obliki je protein TRF2 popolnoma odsoten, kar zaradi genomske nestabilnosti vodi v fuzijo kromosomov, to pa v apoptozo.
Zaključek
S proteini obdana ponavljajoča se šestnukleotidna zaporedja TTAGGG na koncih kromosomov tvorijo telomere, varuhe dednega materiala. TRF2 skupaj s številnimi proteini in gvaninski rep preko specifičnih interakcij omogočajo nastanek zanke in s tem kvartarno strukturo koncev evkariontske DNA. Čeprav gre za zelo stabilno strukturo, pa so skozi celični cikel opazne tri različne oblike.
V prihodnosti lahko pričakujemo še veliko uporabnih odkritij o zgradbi in vlogi telomerov, s katerimi bomo nadgradili dosedanje znanje o njihovi pomembnosti.
Viri
- A. J. Cesare and J. Karlseder, “A three-state model of telomere control over human proliferative boundaries.,” Curr. Opin. Cell Biol., vol. 24, no. 6, pp. 731–8, Dec. 2012.
- D. E. Gomez, R. G. Armando, H. G. Farina, P. L. Menna, C. S. Cerrudo, P. D. Ghiringhelli, and D. F. Alonso, “Telomere structure and telomerase in health and disease (Review),” Int. J. Oncol., vol. 41, no. 5, pp. 1561–1569, Nov. 2012.
- J. Lin, P. Kaur, P. Countryman, P. L. Opresko, and H. Wang, “Unraveling secrets of telomeres: One molecule at a time.,” DNA Repair (Amst)., Mar. 2014.
- T. de Lange, “How telomeres solve the end-protection problem.,” Science, vol. 326, no. 5955, pp. 948–52, Nov. 2009.
- W. Lu, Y. Zhang, D. Liu, Z. Songyang, and M. Wan, “Telomeres-structure, function, and regulation.,” Exp. Cell Res., vol. 319, no. 2, pp. 133–41, Jan. 2013.